IGF family
While we previously discussed the somatomedin hypothesis and the role of IGF-I as it relates to the actions of GH, it has become increasingly clear that the IGF family of growth factors, receptors, and binding proteins are critical players in many specific physiological processes including mammary and ovarian development.
IGF-I and IGF-II are widely expressed endocrine-, autocrine-, or paracrine-acting peptides that regulate cell growth, cell differentiation, maintenance of cell function, and prevention of apoptosis in multiple cell types. Research with IGF-I and IGF-II was initially centered on the somatomedin hypothesis (Hadsell and Bonnette, 2000; Hadsell et al., 2002). Since these early experiments, with the somatomedins now called the IGFs, the view has evolved that the IGFs are also important locally acting autocrine or paracrine stimulators of cell function. The primary signaling receptor for the IGF-I peptide is a tyrosine kinase receptor (IGF-IR) structurally similar to the insulin receptor (see Fig. 12.5). The receptor is widely distributed and is now known to have important roles in normal cell growth and development. Moreover, abnormal stimulation of IGF-IR is implicated in appearance and continuing development of a variety of different types of tumors. In particular, the strongly antiapoptotic activity of the receptor is recognized as being relevant in tumorgenesis. However, other related effects, including effects on cell-cell and cell- extracellular matrix interactions, are also likely important in both normal and abnormal tissue development. Our focus is on normal development, but much of the recent basic work to understand functions of IGFs and IGF-IR is derived from cancer-oriented studies.In the cow, normal circulating concentrations of insulin l-5ng∕mL would have little ability to signal via IGF-IR. IGF-I can also bind to the insulin receptor but with much less affinity, -1000-fold.
The situation is further confounded by the existence of hybrid receptors between IGF-IR and the native insulin receptor (IR) that have higher affinity for IGFs than for insulin. IGF-II binds with high affinity to a receptor that is identical with a receptor for mannose-6-phosphate, but the receptor has no known intracellular signaling function. The affinity of this receptor for IGF-I is about 100-fold lower than for IGF-I and it does not recognize insulin. There are as many as five distinct types of insulin/IGF-I hybrid receptors. The IR isoforms IR-A and IR-B, as well as IGF-IR, have the ability to form both homo- and heterotetramers. This clearly can change the diversity of signaling induced by binding of insulin or IGFs. Signaling pathways and biological effects are best characterized for IGF-IR and IR-B, but experiments in cell culture model systems support the idea that combinations of native and hybrid receptors allow both overlapping and unique physiological effects. In general, stimulation of either IGF-IR or IR-A is associated with cell cycle progression but stimulation of IR-B is more closely related with metabolic events. These major properties were illustrated in elegant molecular studies in which the cytoplasmic domains for IR-B and IGF-IR receptors were swapped. In the normal situation, there are overlapping as well as specific signaling events associated with activation of IR-B or IGF-IR (Hadsell and Bonnette, 2000).Studies with rodents in which various elements of the IGF-I axis were deleted have confirmed that IGF-I and/or IGFI-R are essential for normal mammary development (Kleinberg et al., 2000). Since animals homozygous for absence of the IGF-IR do not survive after parturition, these experiments required the transplantation of the fetal mammary analogue from approximately day-18 fetuses into the cleared mammary fat pads of syngeneic hosts. Growth of rudimentary mammary structures from IGF-IR null mice, compared with tissue from wild-type, was minimal.
In mice without expression of IGF-I, the development of terminal end buds required replacement with IGF-I. Neither estradiol or GH alone (classic mammary acting hormones) nor the combination had any effect on prepubertal mammary development in these knockout mice. These and related rodent experiments show that in normal peripubertal mammary development, GH acts to bind to GH receptors in the stromal tissue. This is associated with local production of IGF-I, which in turn promotes the development of the mammary ducts. Certainly, overexpression of recombinant IGF-I in the mammary glands of transgenic mice promotes premature development of alveolar buds in prepubertal animals (Weber et al., 1998).In addition to IGF-I and IGF-II, there are six IGF- binding proteins (IGFBPs) and nine related proteins (IGFBP-rP) that affect the actions of IGF-I and II. The IGFBPs are well characterized and bind IGF-I with ~10-fold higher affinity than the IGFBP-rPs. The IGFBPs have several functions, including prolonging the half-life of IGF-I, transporting IGF-I from the circulation, and localizing IGF-I to potential target cells (Clemmons, 1998; Duan, 2002). Locally produced IGFBPs provide a mechanism to target or localize IGFs within particular tissues or cells and thereby alter biological responses to IGFs.
It is worth taking a moment to discuss this idea of binding proteins and how they can be studied. A typical procedure is to separate solutions of proteins suspected to contain binding proteins by standard SDS-PAGE electrophoresis. These proteins that are then separated in columns or lanes based on molecular weight and/or charge and are then transferred to a membrane (essentially like a sheet of paper). This membrane is subsequently washed in an appropriate buffer, then incubated in a solution containing I125 radiolabeled IGF-I or IGF-IL The iodinated protein is allowed to bind, then the membrane is washed and exposed to X-ray film. The position of proteins that bind to the IGF then appears as black, dense bands on the developed film.
These bands can then be photographed and/or scanned and evaluated for the level of density. When specific antibodies to the binding proteins for a particular species are available, these ligand blots can be supplemented using western blotting. Figure 12.37 gives an example of an IGF-I ligand blot-binding protein prepared from conditioned medium collected from bovine mammary epithelial cells. In this example, there is abundant expression of IGFBP-2 and IGFBP-3 (as confirmed by western blotting) and a lower molecular weight binding protein that is probably IGFBP-4 based on molecular weight (Romagnolo et al., 1994). The secretion of these binding proteins is increased when the cells are incubated with native IGF-I or two analogs of IGF-I that exhibit reduced capacity to interact with binding proteins.Along with locally produced IGF-I made by epithelial cells (see Fig. 12.18), as illustrated by experiments with bovine mammary tissue (Weber et al., 2000), mammary cells also synthesize several IGFBPs, including IGFBP-I, 2, 3, and 5. IGFBP-3 is usually touted as an inhibitor of responses to IGF-L For example, Figure 12.38 shows increased proliferation of mammary tissue following treatment with estrogen, GH, or a combination of the two, and the associated mirror image a decrease in tissue concentrations of IGFBP-3. This suggests that at least some of the proliferation induced with these classic mammogenic hormones is mediated by a local reduction in mammary tissue
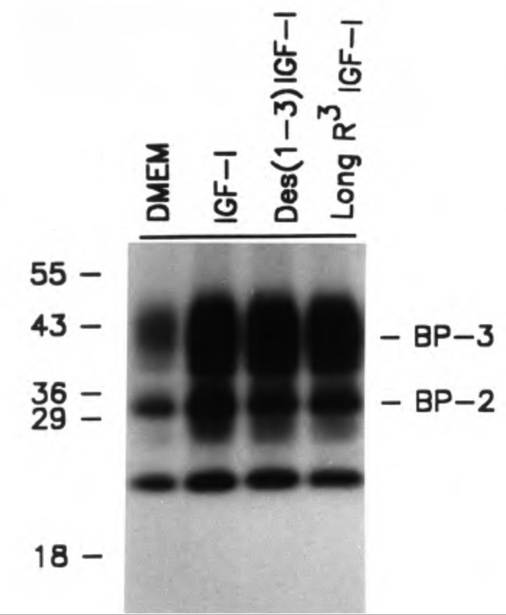
Fig. 12.37. IGF-l-bincling proteins. Effects of incubation of bovine mammary cells with IGF-I or selected IGF-I analogs are shown. Adapted from Romagnolo et al. (1994).
IGFBP-3. There is also increased tissue IGF-I, again suggesting the importance of IGF-I axis molecules in regulation of mammary development (Berry et al., 2001).
In addition to their effects related to mammary development and function in farm animals, the IGFs and binding proteins are also critical in ovarian development.
In fact, in all mammalian species studied, IGF-I stimulates the granulosa cell growth and steroidogenesis. However, changes in local concentrations of IGFs are not as critical as marked alterations in the tissue concentrations of IGFBPs. These changes are responsible for enhanced availability of free IGF-I during terminal follicular growth, then increased synthesis of IGFBP-2, 4, and 5 to promote atresia. Specifically, as reviewed by Monget et al. (2002), in the ewe, sow, cow, and mare, Intrafollicular concentrations of IGFBP-2, -4, are markedly decreased from the 1- to 2-mm follicles to the larger preovulatory follicles. By contrast, Intrafollicular concentrations of these same IGFBPs as well as IGFBP-5 in the ruminants greatly increase in atretic follicles. This pattern of change is due to two events: variation in mRNA expression and selected degradation caused by expression of intra- follicular proteases. These examples simply illustrate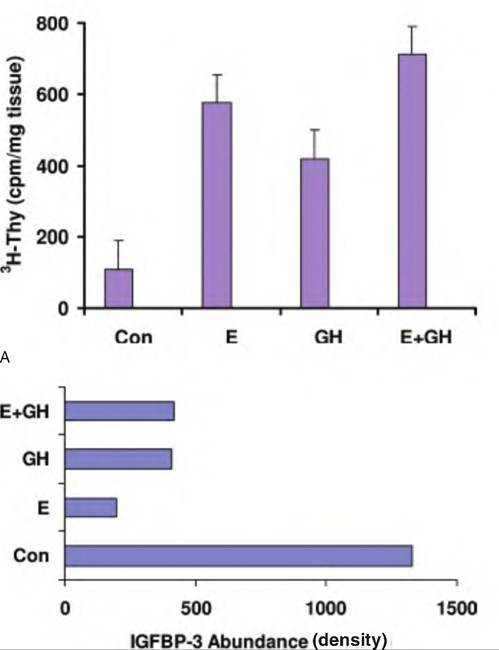
B
Fig. 12.38. Thymicline incorporation in mammary tissue and IGFBP-3. Panel A shows increased DNA synthesis in mammary tissue of heifers treated with placebo (Con), estradiol (E), growth hormone (GH), or both (E + GH). Panel B illustrates the relative abundance of IGFBP-3 in mammary tissue from these same animals. Note that increased proliferation appears negatively correlated with decreased tissue IGFBP-3. Adapted from Berry et al. (2001).
two areas of tissue and cell development that are impacted by this important family of growth factors.