EGF family
Epidermal growth factor (EGF) was discovered by accident in the 1960s when unexpected biological activity (not attributed to nerve growth factor) occurred following injections of extracts of murine salivary glands into test animals.
These effects were associated with precocious eyelid opening and tooth eruption in neonates. Subsequent studies showed that preparations of EGF stimulated the proliferation of isolated epidermal cells and epidermal tissue explants, hence the name EGE Study of a human epidermal cancer cell line (A-431) was fundamental in subsequent studies because these mutated cells greatly overexpressed receptors for EGF (~3 ? IO6 receptors per cell). This property allowed for the isolation and structural characterization of the receptor. The availability of relatively large quantities of the receptor also led to the discovery that the addition of EGF to isolated membranes stimulated the phosphorylation of both endogenous membrane proteins as well as many exogenously added proteins. It was subsequently shown that the receptor protein was a 170 kDa transmembrane glycoprotein whose external domain formed the binding site for EGF and whose cytoplasmic domain possessed tyrosine kinase activity. EGF bound to the external portion of the receptor activates the cytoplasmic or catalytic domain of the receptor to produce autophosphorylation and subsequently phosphorylation of cytoplasmic substrates essential for EGF action. These observations were important because they were among the first to directly link ligand binding, receptor activation, and phosphorylation as general mechanisms of action for many growth factors.This family of proteins contains at least ten members including: EGF, heparin-binding EGF, transforming growth factor a (TGF-θc), amphiregulin, neuroregulins (four subtypes), and several heregulins.
Each of these structurally similar proteins acts by binding to one of several related membrane receptors called type I receptor tyrosine kinases (RTKs) or the ErbB family of receptors. The usual EGF receptor is called ErbB-I, but at least four variants are known. Certain receptors in the family, ErbB-2 (also called HER2 or Neu), contribute to the aggressive phenotype of some human breast carcinomas and related poor prognosis. A homolog of EGF, TGFoc, was first isolated from the medium of oncogene-transformed cells and a transforming avian retrovirus subsequently was shown to code for synthesis of an abbreviated form of the EGF receptor. EGF and TGFoc are highly expressed in early embryonic development. Amphiregulin and HB-EGF are secreted heparin-binding growth factors. The ErbB family of receptor kinases is essential for development of the nervous, cardiovascular, gastrointestinal, and other systems.Dysfunction of these receptors is common in many forms of carcinogenesis, which explains much of the emphasis for work in this area. A direct connection between the ErbB family and cancer was established with the breakthrough that the retroviral oncogene V-ErbB, which is transduced by the avian erythroblastosis virus, is a form of an avian EGF receptor. This then led to subsequent discoveries showing that overexpression of ErbB agonist proteins and/or overexpression or inappropriate expression of ErbB receptors were important in a variety of human cancers. In fact, a humanized monoclonal antibody against ErbB-2 called Herceptin was the first such product approved for human use. It was designed specifically for breast cancer patients with tumors that overexpress ErbB-2 (Harris et al., 2003; Stern, 2003).
EGF is important in normal mammary gland development in mice and likely other species as well. In particular, there is compelling evidence to suggest that estrogen (a potent mammogenic hormone from the ovary) likely induces local production of EGF agonists (EGF itself, TGFoc, or amphiregulin) and that these agents are important in alveolar development.
Receptors for EGF are present in regions of rapidly growing mammary ducts, including the surrounding stromal cells. The importance of these proteins in mammogen- esis in farm animals is less well established but specific EGF receptors are found in mammary tissue from sheep and cows (Plaut, 1993) and expression of EGF mRNA has been shown for bovine mammary tissue (Koff and Plaut, 1995). Ligand-binding assays using either radiolabeled EGF or TGFoc show a single class of high-affinity-binding sites in mammary tissue of sheep and cows during gestation and into lactation. However, the number of receptors is greater during mid than late pregnancy or during lactation. Expression of TGF-α occurs in the bovine and amphiregulin occurs in ovine mammary tissue. Addition of either TGF-Ot or EGF stimulates DNA synthesis in explants from midpregnant heifers, in epithelial cells from heifers or pregnant cows and sheep. Sheffield et al. (1998) studied the effect of EGF on DNA synthesis in mammary explants prepared from tissue taken from midpregnant heifers, either before or after xenotransplantation into nude mice. For freshly prepared explants, concentrations of EGF of less than IOnM only stimulated DNA synthesis after 2 days in culture. For Xenotransplanted tissues, priming of mice with estrogen and progesterone for 10 days increased the sensitivity of the tissue to EGF when explants were removed and tested in culture. Failure of EGF to stimulate DNA synthesis in mice given hydrocortisone and Prl suggests that differentiation of the alveolar cells is incompatible with EGF-induced growth. Indeed, we have noted that EGF can also mimic lactogenic effects of Prl or hGH on secretion of caseins by mammary explants taken from mid-gestation heifers but not secretion of the milk protein oc-lactalbumin. This indicates that the role of EGF in cell proliferation and differentiation in the ruminant mammary gland is complex. EGF may stimulate cell proliferation in some situations but impact cell differentiation (milk component synthesis/secretion in this case) in others. Interrelated actions with other mammary active hormones and growth factors is likely. Some of the effects of EGF on bovine mammary tissue are illustrated in Figure 12.39.For tissues that synthesize EGF or its related ligands, these growth factors are made as transmembrane glycoproteins. The EGF-Iike sequences are external to the
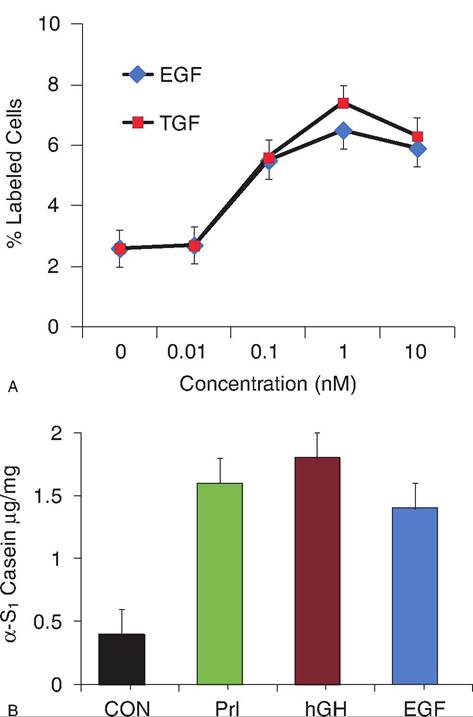
Fig. 12.39. Panel A illustrates a concentration depending increase in mammary cell proliferation (% epithelial cell nuclei labeled with tritiated thymidine). Panel B illustrates that addition of EGF can have lactogenic effects on bovine mammary tissue as evidenced by an increase the secretion of the milk protein (α-S1 casein) by mammary explants from mid-gestation heifer culture. This lactogenic response is similar to responses following addition of bovine prolactin or human growth hormone. Interestingly, addition of EGF did not increase secretion of the milk protein Oi-Iactalbumin as did addition of both bPrl and hGH.
plasma membrane. Requirements for secretion are not completely understood but involve the action of a protease that cleaves the membrane-bound protein for release of the EGF (or relative) into the interstitial fluid. Induced overexpression of TGF-Ot under control of mammary promoters in rodents leads to the appearance of mammary tumors. This fits with measurements showing that some spontaneous human mammary tumors also express large amounts of TGF-OC or EGF receptor. It is also important to remember that responses of cells or tissues in culture may not necessarily reflect the response of tissue in the complex environment of the intact animal. Presence of EGF receptors and ligands and demonstrated response of ruminant mammary cells to EGF support a role for these growth factors in mammogenesis, but much more information is needed to determine exact roles. In short, it is not known if the EGF family of proteins and receptors is largely permissive or if they have an essential direct impact in ruminant mammary development.
The EGF family is also important in the growth and development of hair, so there is much interest in impacts of these growth factors on wool and other animal hair products. Specifically, the hair follicle has two closely aligned epithelial cell layers. The inner root sheath cells produce TGF-α but do not express EGF receptor (ErbB-I), but the opposite is true for the outer root sheath (receptors but no TGF-α). For normal growth it is essential that newly formed TGF-OC be processed for release and then rapidly bound by receptors in the outer sheath of cells. In the case of EGFR- or TGF-0C-null mice, which are engineered without the receptor and/or ligand, hair follicle formation is disturbed. When this local TGF-oc escapes capture, it acts an attractor of other cells into the region of the hair follicle so that the hair coat is abnormal and whiskers are wavy (Mann et al., 1993).
More on the topic EGF family:
- EGF family
- Other hormones and growth factors
- Contents
- TGF-β family
- Akers R. Michael, Denbow D. Michael. Anatomy and Physiology of Domestic Animals. 2nd edition. — Wiley-Blackwell,2013. — 685 p., 2013
- 47 Cancer of the Uterine Corpus