Immunity and Defense
Christina M. Jeffries, AAS, CVT and Morgan Rodgers, BS, AAS, CVT
OUTLINE
INTRODUCTION, 318
ANATOMIC ORGANIZATION OF THE
IMMUNE SYSTEM, 318
Skin and Mucous Membranes—The First Line of Defense, 318
Organs and Tissues—Internal Protection, 318 Lymphatic System, 319
Red Bone Marrow, 320
FUNCTIONAL ORGANIZATION OF THE
IMMUNE SYSTEM, 320
Innate Immune System, 321
Adaptive (Acquired) Immune System, 329
Active Immunity, 334
Passive Immunity, 335
Mechanisms of Disease, 335
Hypersensitivity Reactions, 336
LEARNING OBJECTIVES
When you have completed this chapter you will be able to:
1.
List the organization and functions of the immune system. 2. Describe the structures and functions of the immune system, including the white blood cells, lymph nodes, spleen, thymus, tonsils, and GALT.
3. Differentiate between internal and external innate immune functions.
4. Understand complement proteins, cytokines, and interleukins along with their functions.
5. Differentiate between specific and nonspecific immune reactions.
6. Describe various nonspecific immune activities.
7. Differentiate between and understand the components of cell-mediated versus humoral immunity.
8. Describe the classes of immunoglobulin.
9. Differentiate between active and passive immunity.
10. Understand what can occur when the immune system doesn't function properly.
VOCABULARY FUNDAMENTALS
Active immunity ahck-tihv ihm-myoo-nih-te
Adaptive immunity ah-dahp-tihv ihm-myoo-nih-te
Antibody ahn-te-boh-de
Antigen ahn-teh-jehn
Apoptosis ahp-ohp-to-sihs
Complement kohm-pleh-mehnt
Cytokine sι-to-kιn
Disease dihz-ez
Fever fe-var
Infection ihn-fehck-shuhn
Inflammation ihn-fluh-ma-shuhn
Innate immunity ihn-at ihm-myoo-nih-te
Interferon ihn-tar-feer-ohn
Macrophage mah-kro-faj
Memory cell mehm-ohr-e sehl
Microbe mι-krob
Passive immunity pah-sihv ihm-myoo- nih-te
Pathogen pahth-o-jehn
Pathogenicity pahth-o-jeh-nihs-ih-te
Phagocyte fahg-o-sιt
Phagocytosis fahg-o-sι-to-sihs
Virulence vihr-u-luhnz
Virulence factor vihr-u-luhnz fahck-tar
INTRODUCTION
Every day an animal's body encounters millions of microorganisms.
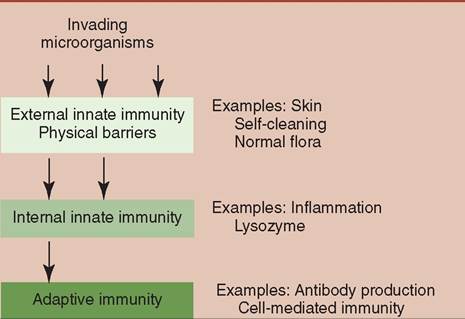
They are looking for food and a warm place to live and breed. Although many are harmless, others are capable of causing illness or even death. Thankfully, the body has the ability through its immune system to defend itself against pathogens (organisms capable of causing disease). The immune system has evolved to act as the security system of the body responsible for recognizing foreign material and protecting the body from anything that is not part of the animal. Its major function is distinguishing "self" from "nonself" cells, allowing easy identification and destruction of potential pathogens. Without the immune system, animals would be in a constant state of disease, if they survived at all. The first line of defense of the immune system is made of external barriers. The second line of defense includes cellular and chemical components. Together they make up the innate immune system. If a pathogen evades the first and second lines of defense, the third line of defense, also known as the adaptive immune system, will be activated (Figure 13-1). ![]()
FIGURE 13-1 The three major barriers that protect an animal's body against microbial invasion. Each barrier provides a more effective defense than the previous one. (Redrawn from Tizard I: Veterinary immunology, ed 9, St Louis, 2013, Saunders.)
ANATOMIC ORGANIZATION OF THE IMMUNE SYSTEM
SKIN AND MUCOUS MEMBRANES— THE FIRST LINE OF DEFENSE
The body's first line of defense against potential pathogens consists of physical barriers that prevent the pathogens from entering the body. By limiting pathogen entry, the body minimizes the effort required to defend the animal. If pathogens cannot enter, they cannot cause disease.
The first, and in many ways, most important defense mechanism is the skin. Covering the entire body surface, the skin serves as a physical barrier that protects the vital organs inside.
In addition to serving as a physical barrier, the surface of the skin supports a variety of resident microorganisms (normal flora) that recognize and destroy potential invaders. Unless there is a break in the skin, most pathogens are destroyed before they can enter the body. Additionally, the acidic pH and fatty acid content of sweat inhibit bacterial growth. Of course, if the animal's body were completely sealed off from the environment, it would be unable to acquire food, oxygen, and water—all requirements for life. It would also be unable to remove the waste products of metabolism. One or the other would eventually kill the animal. To sustain life, there must be pathways through the protective layer of the skin. The largest of these pathways include the respiratory, gastrointestinal, and urinary tracts. Although these openings are required, to leave them completely open would be just as dangerous as having no protective barrier. Thus, the body has several protective mechanisms that allow the materials entry into these pathways while still protecting the animal. For example, the respiratory system uses a combination of cilia and mucus to trap and remove potential pathogens away from the lower respiratory tract. The digestive system uses the acidity of the stomach to kill many microbes that gain entry through ingestion. In addition, fluids such as tears, saliva, nasal discharges, and urine flush pathogens from the body.
ORGANS AND TISSUES— INTERNAL PROTECTION
SPLEEN
If a microbe is able to penetrate these physical and chemical barriers, there are internal organs with immunologic functions. The largest of these organs is the spleen, which is composed of white pulp and red pulp. White pulp is made of lymphoid tissue and is the portion of the spleen with immunologic functions. It surrounds blood vessels and contains phagocytic cells that react to antigens in the bloodstream. One of the functions of these cells is removing the antigens from the blood via phagocytosis, and initiating an immune response to produce antibodies against the antigen.
Recall from Chapter 12 that the spleen acts as a reservoir for additional blood supply in the red pulp. Specialized blood- filled sinuses make up the red pulp, which is also responsible for removing worn, damaged, or aged blood cells by the action of tissue macrophages. Animals are capable of living without the spleen, and after a splenectomy, macrophages and lymphoid tissue in other areas of the body are able to compensate for the loss. LYMPHATIC SYSTEM
The next largest component of the immune system is the lymphatic system. The lymphatic system is responsible for collecting and returning excess interstitial tissue fluid to the cardiovascular system. Lymph vessels parallel the vessels of the circulatory system. As blood flows through arteries and capillaries, some of this fluid seeps into the interstitial tissue space. Lymphatic capillaries drain any excess fluid, as lymph, into lymphatic vessels (see Figure 12-23). As lymph travels through lymphatic vessels, it picks up water, electrolytes, sugar, and lymphocytes. Lymph collected from the digestive system is called chyle, which often contains small fat molecules called chylomicrons (which are responsible for the postprandial lipemia found in blood samples drawn just after an animal has eaten). Lymph travels through the vessels of the lymphatic system to the thoracic duct, which empties into the systemic circulation in the area of the right atrium of the heart. This mixes the lymph with blood returning to the heart. In this way the lymphatic system also helps maintain osmotic pressure and fluid balance.
LYMPH NODES
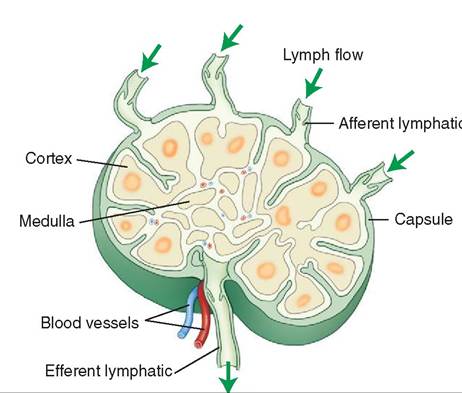
Located throughout the body are lymph nodes, small structures that are responsible for lymph filtration as it travels back to the systemic circulation. Lymph enters the cortex of the lymph node via afferent vessels in most species (except pigs). This is where clusters of lymphocyte nodules exist. The lymph then passes into the medulla of the lymph node, which contains macrophages that remove potential pathogens such as microorganisms, cancer cells, or foreign debris.
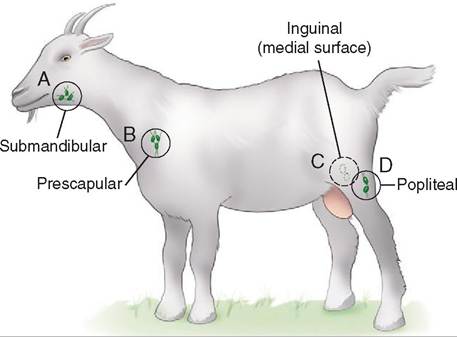
After passing through the medulla, the lymph exits the lymph node via the efferent vessels. As it moves through the lymph node, some of the lymphocytes enter the lymph and travel to other areas of the body, where they are capable of mounting an immune response (Figure 13-2). As lymph returns from the periphery (e.g., the limbs), it has to flow through at least one lymph node. Although there are lymph nodes dispersed throughout the body, the peripheral lymph nodes can easily be palpated, and thus are easiest to assess. The peripheral nodes include the submandibular (caudal to the mandible), prescapular (cranial to the shoulder), axillary (where the front limb joins the trunk), inguinal (near the groin), and popliteal (distal/caudal aspect of the hamstring muscles) (Figure 13-3). As lymph fluid passes through successive nodes, the likelihood that any potential pathogen will survive decreases significantly.
Lymph from specific areas of the body will always pass through the same lymph node(s), which may aid in determining the location of an inflammatory response, infection, or tumor. A lymph node near an affected area becomes enlarged because of increased lymphocyte action (multiplication) and macrophage accumulation in response to the presence of an antigen. For this reason, in cases of suspected cancer it is common to biopsy the lymph nodes that filter lymph from an area where the tumors are present. Evaluation of the biopsy can help determine whether the cancer has begun to metastasize (spread) and assist in planning treatment protocols. If the cancer has metastasized to regional lymph nodes, they may need to be removed along with the original tumor.
MALT
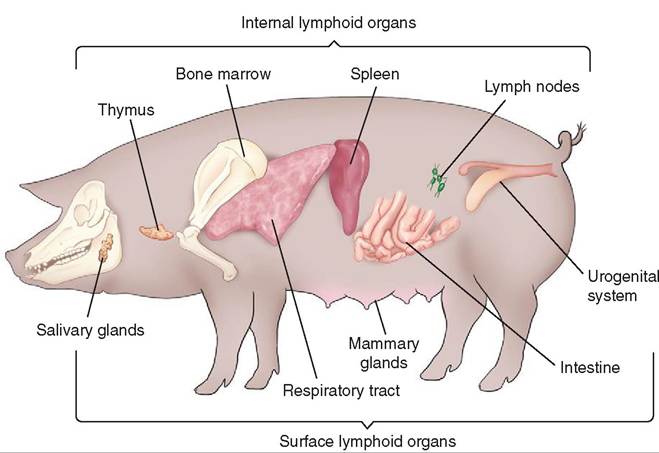
In addition to the peripheral lymph nodes, there are clusters of lymphoid tissue in various areas throughout the animal's body (Figure 13-4). Referred to as mucosa-associated lymphatic tissue (MALT), these small lymphatic nodules are located near mucosal surfaces but are not encapsulated like a lymph node.
The function of MALT is to identify antigens and mount an immune response against them. There are subcategories of MALT that include conjunctiva-associated lymphoid tissue (CALT), nose or nasopharynx-associated lymphoid tissue (NALT), and gut-associated lymphoid tissue (GALT). The function of GALT is to ensure that pathogens that survive the acidic environment of the stomach cannot infect the animal via the gastrointestinal (GI) tract. ![]()
Lymph flow
FIGURE 13-2 The major structural features of a typical lymph node.
![]()
FIGURE 13-3 Locations of common palpable lymph nodes in a goat. A, Submandibular lymph node; B, prescapular lymph node; C, Inguinal lymph node (in the groin area); D, popliteal lymph node.
![]()
FIGURE 13-4 The major lymphoid tissues of the pig, a typical mammal.
TONSILS
Tonsils are found in the epithelial tissue in the pharynx, larynx, and urinary and reproductive tracts. They are part of the MALT system. Tonsils house lymphocytes to destroy foreign material before it is able to cause disease. Tonsils found in the posterior nasal cavity are called adenoid (pharyngeal) tonsils, whereas tonsils found in the posterior oral cavity are called palatine tonsils. Unlike lymph nodes, which are located along lymph vessels, tonsils are present at the beginning of the lymph drainage system and lack a capsule. Tonsils are also found in the prepuce and vagina where they perform a similar function.
PEYER'S PATCHES
Peyer's patches are aggregations of lymphoid tissue in the small intestine of animals such as cattle, sheep, pigs, horses, and dogs. The majority of Peyer's patches are found in the lining of the ileum, the final section of the small intestine. A smaller percentage of Peyer's patches are found in the jejunum (the middle section of the small intestine).
THYMUS
The thymus is found in young animals as an additional concentration of lymphoid tissue located in the mediastinum. The thymus is where T lymphocytes mature before they migrate into other lymphoid tissues and blood, and where T cells are programmed to fight specific antigens. The cell lines for the T lymphocytes are established while the animal is a juvenile. As the animal matures, the thymus atrophies and it is usually undetectable in an adult animal. However, microscopic clusters of cells in the area continue to produce T cells throughout the life of the animal.
RED BONE MARROW
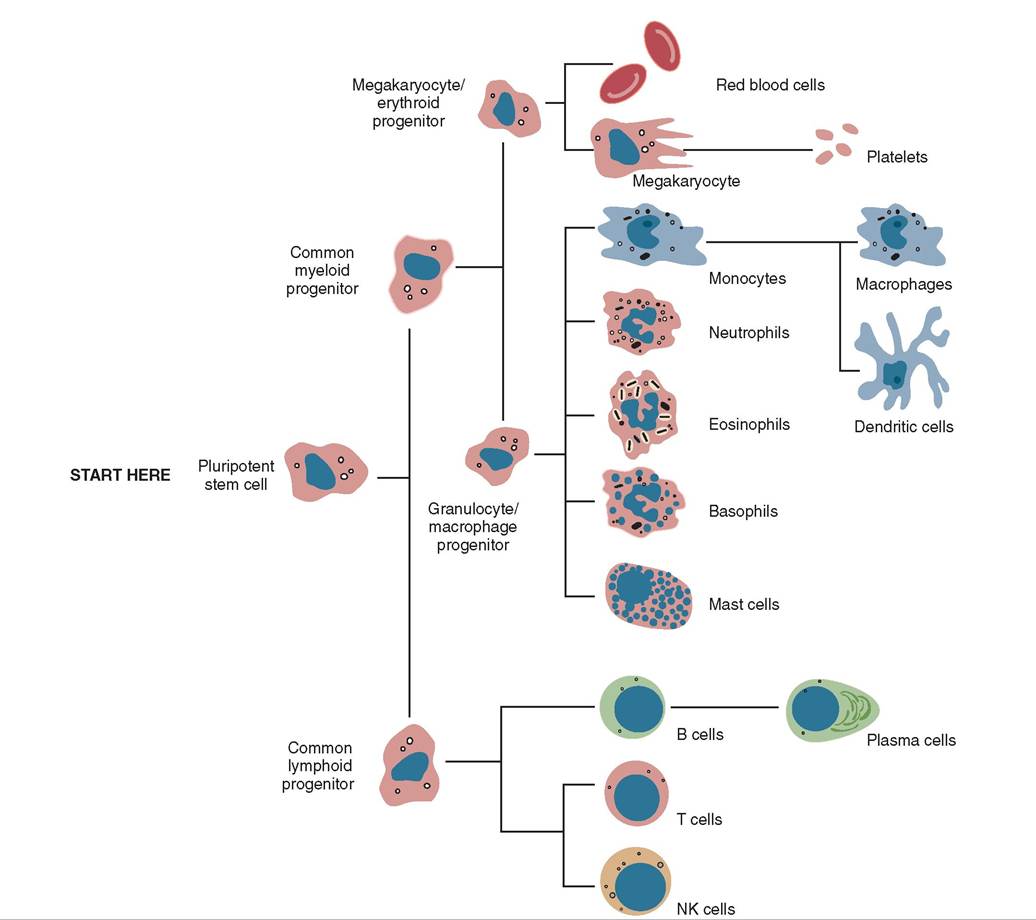
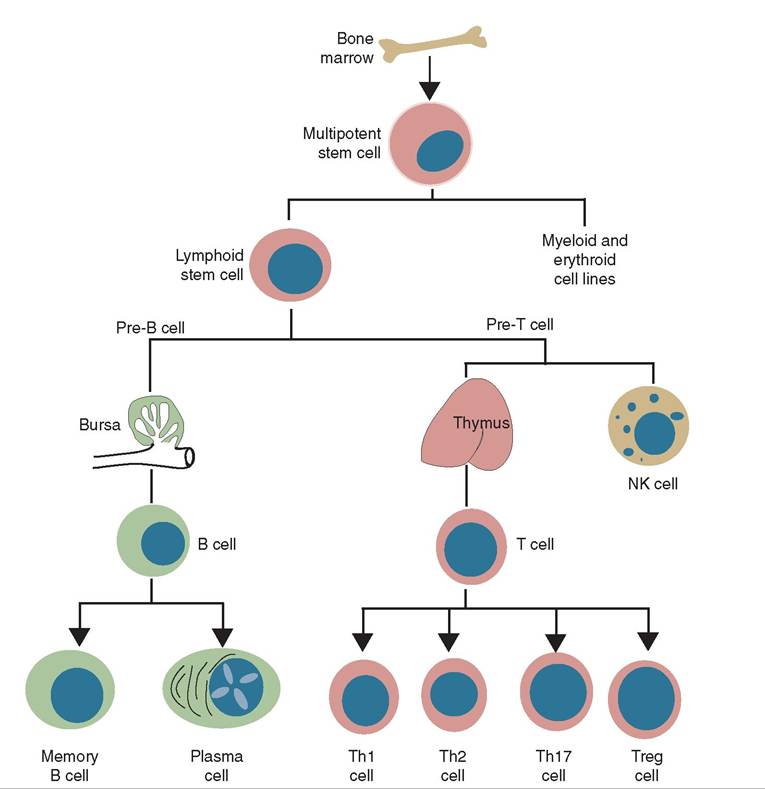
Red bone marrow, although not strictly an immune organ, is responsible for the production of all white blood cells, the soldiers of the immune response. Look at Figure 13-5. Notice that the pluripotent stem cell will become either a common lymphoid progenitor or a common myeloid progenitor. The common lymphoid progenitor leaves the bone marrow and matures to one of the various types of lymphocyte in lymph tissue in other parts of the body. The common myeloid progenitor stays in the bone marrow and the cells that develop from it will mature in the bone marrow.
When the monocyte is called into tissue it becomes a macrophage. Depending on the location in which it is found, it can have a specific name. For example, a macrophage in the liver is called a Kupffer cell, in the central nervous system it is a microglial cell, in bone and bone marrow it is an osteoclast, and in the epidermis and lymph nodes it is a dendritic cell. The dendritic cell is a sentinel macrophage that can capture invading pathogens and take them to a lymph node for destruction. We'll discuss the importance of dendritic cells later in the chapter. See Chapter 12 for more detailed descriptions of the different types of white blood cell.
TEST YOURSELF 13-1
1. What is the main function of the immune system?
2. What organs are involved in immunity?
3. Describe how the lymphatic system protects the body from disease.
4. Where is MALT found?
5. What is the significance of the thymus?
FUNCTIONAL ORGANIZATION OF THE IMMUNE SYSTEM
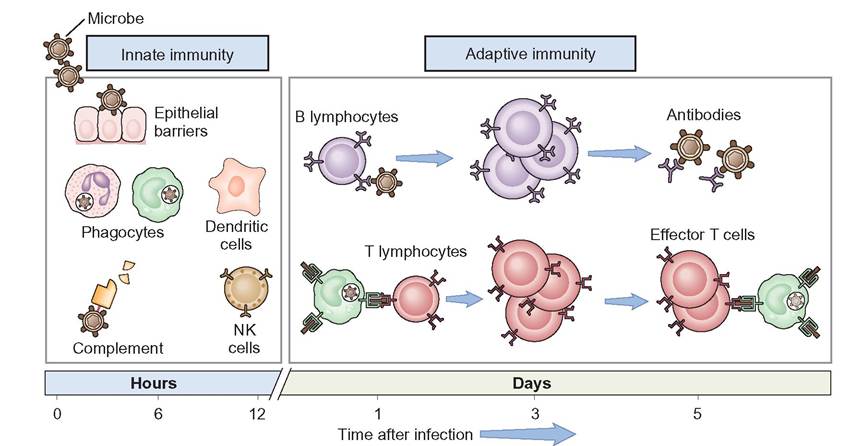
The immune system is functionally divided into two categories: innate and adaptive immunity (Figure 13-6). The innate
![]()
FIGURE 13-5 The origin of cells from the red bone marrow. (From Tizard I: Veterinary immunology, ed 9, St Louis, 2013, Saunders.)
immune system is rapid, nonspecific and destroys “nonself” invaders indiscriminately. It is present at birth and uses physical, chemical, and cellular components to protect the body from anything identified as “nonself.” The innate immune system is unable to target specific organisms; instead, it destroys all “nonself” invading organisms by the same mechanisms.
The adaptive immune system targets specific organisms, but it is slower to respond to an invading organism. It is not present at birth but develops and adapts as the animal matures and is exposed to a variety of antigens. Once an animal is exposed to an antigen, the adaptive immune system uses antibodies, memory cells, plasma cells, B lymphocytes, and T lymphocytes provide immunity.
TEST YOURSELF 13-2
1. What are the two main subcategories of the immune system?
2. How does specific immunity differ from nonspecific immunity?
3. Adaptive immunity is nonspecific immunity. True or False?
INNATE IMMUNE SYSTEM
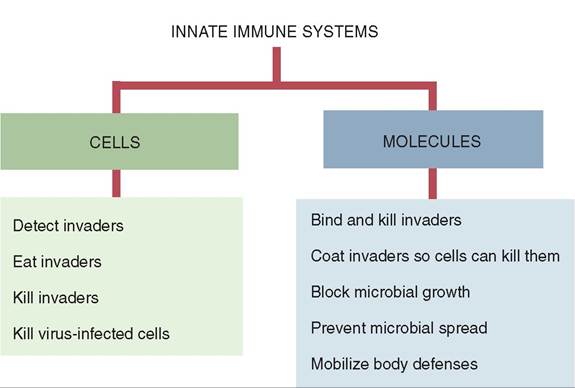
The innate or nonspecific immune system works independently or in conjunction with the adaptive immune system to prevent disease by providing mechanical (anatomic) and cellular barriers (Figure 13-7). It is present at
![]()
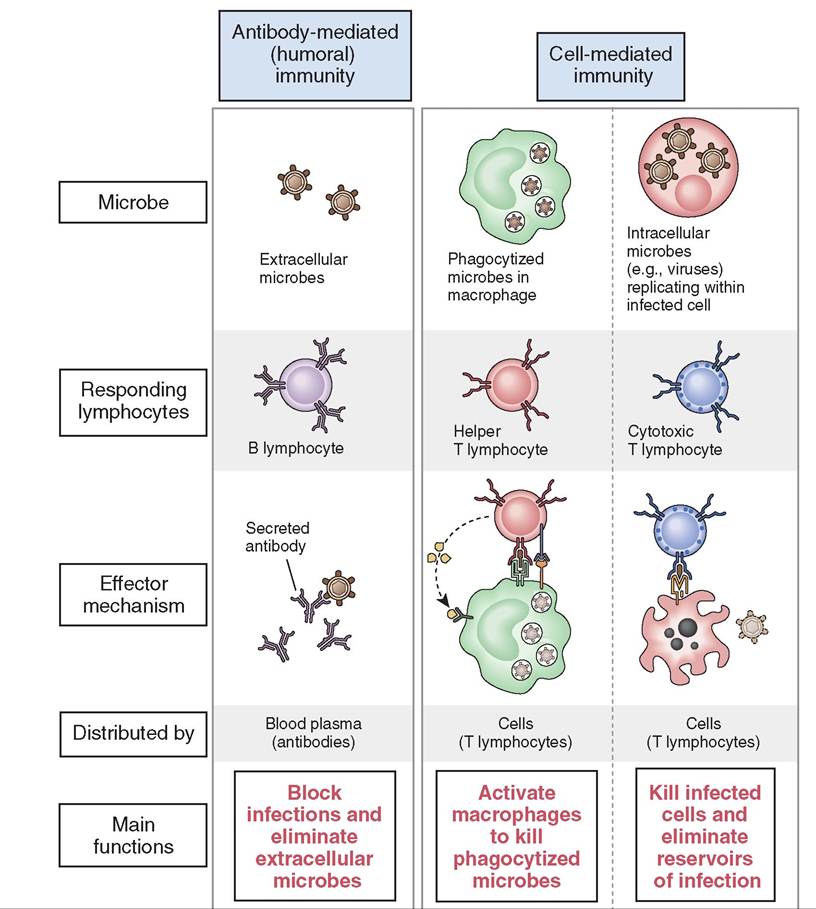
FIGURE 13-6 The principal mechanisms of innate and adaptive immunity. The mechanisms of innate immunity provide the initial defense against infections. Some of the mechanisms prevent infections and others eliminate microbes. Adaptive immune responses develop later and are mediated by lymphocytes and their products. Antibodies block infections and eliminate microbes, and T lymphocytes eradicate intracellular microbes. NK, Natural killer cell. (From Abbas AK: Basic immunology updated edition: functions and disorders of the immune system, ed 3, St Louis, 2010, Saunders.)
![]()
FIGURE 13-7 The innate immune system consists of a collection of multiple subsystems. They can be divided into the cells that largely eat and kill invaders and the molecules that bind and kill the invaders. (From Tizard I: Veterinary immunology, ed 9, St Louis, 2013, Saunders.)
birth and remains virtually unchanged during the life of the animal.
Innate immunity acts as the animal's first and second lines of defense against pathogens that could cause disease in an animal. The innate immune system does not recognize specific pathogens, rather macrophages recognize common structures shared by large groups of pathogens. These pathogen-associated molecular patterns (PAMPs) are found omnbrtahnee me surface of invading pathogens. The
membrane surface of a macrophage or dendritic cell has receptors called pattern-recognition receptors (PRRs) that fit the PAMP on the pathogen's cell membrane. This allows the macrophage to recognize and attach to the pathogen and trigger the innate immune system. Normal, nonpathogenic microbes (normal flora) are recognized as self and will not trigger the innate immune system.
EXTERNAL INNATE IMMUNITY—THE FIRST LINE OF DEFENSE
Anatomic barriers that include structures on the surface of the body that prevent disease provide external innate immunity. ⅞e skin, the largest organ in the body, has a thick layer of kn’zti^ed epithelial tissue that is impermeable to a majority of pathogens. The keratinized epithelial cells have antimicrobial properties that inhibit bacterial growth. When the layer of dead keratinized cells is sloughed off the skin it takes with it any microorganisms that were clinging to the cells. In order for this to be an effective means of disease prevention, the skin must be intact to prevent pathogens from entering the body. When imperfections such as small cuts or lacerations are present on external barriers, pathogens can invade the body tissue and circulate to other areas of the body. Therefore, broken skin is always a weaker defense than intact tissue.
Mucous membranes of the epithelium that line the respiratory, digestive, urinary, and reproductive systems also have unique structures that provide innate immunity. Mucus produced by mucous membranes can trap pathogens. Cilia on the borders of epithelial cells move the pathogens away from entry into the body. For example, in the upper respiratory tract, mucus and cilia trap and propel foreign materials away from the lungs, preventing entrance into the lower respiratory tract. This is important because the lower respiratory tract is an ideal moist and warm location for bacterial growth.
Tears, saliva, and nasal discharge production create a flushing action that helps prevent infection in the eyes, mouth, and nose. Additionally, they contain lysozyme and phospholipase that break down bacterial cell walls and membranes, preventing microbial growth in these regions. The acidic environment in the stomach kills many of the microbes in the food that is ingested. If pathogens penetrate through these external barriers and into deeper tissues, the second line of defense is activated.
INTERNAL INNATE DEFENSE—THE SECOND LINE OF DEFENSE
Once a pathogen has made its way past the physical barriers of the innate immune system, the body tries to control the spread of the infection through acute inflammation. Phagocytes, natural killer cells, interferons, complement receptors, and PRRs play a key role in the inflammatory response.
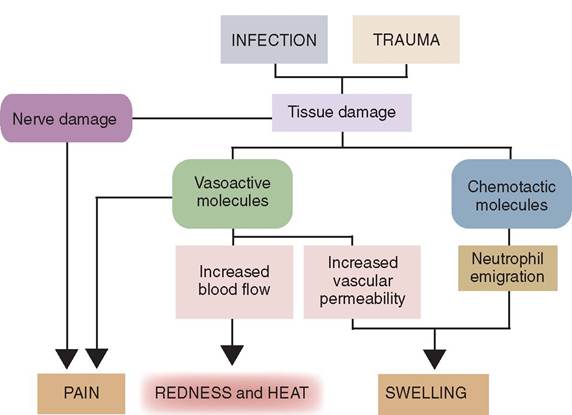
INFLAMMATION. Inflammation is the body's localized reaction created by the innate immune system in response to trauma, infection, chemical exposure, or excessive heat. The four cardinal signs of inflammation are: redness, swelling, heat, and pain g Figure 13-8). Loss of function is another attribute of inflammation that is sometimes considered the fifth sign.
During pathogen invasion, tissue cells are damaged. This triggers the release of various chemicals from specific cells, such as mast cells, that mediate the inflammatory response and result in the signs listed above. These chemicals include histamine, prostaglandins, leukotrienes, and cytokines.
During the inflammatory response arterioles dilate and venules constrict, which leads to increased blood flow and increased capillary permeability at the injured site. This allows large numbers of phagocytic white blood cells (WBCs) to enter the injured site and ingest foreign or cellular debris. Once the pathogens and/or foreign debris have been eliminated, the tissue begins to heal and inflammation is reduced.
Fever represents a systemic inflammatory response where the chemical mediators are carried throughout the body. An elevated body temperature can prevent proliferation of many microbes by creating an environment that exceeds their optimum temperature for growth. This prevents further pathogen growth and occasionally causes pathogen death. A fever can also result in increased cellular metabolism that in turn may cause more rapid phagocytosis, an acceleration of lymphocyte production, and increased antibody production.
Fever has many benefits to the immune system; however, a significant fever can be harmful to the body. An excessively high temperature (>~104° F (40° C)) may cause proteins to denature in the body and can lead to a number of problems,
![]()
FIGURE 13-8 The major signs of acute inflammation and how they are generated. (From Tizard I: Veterinary immunology, ed 9, St Louis, 2013, Saunders.)
CLINICAL APPLICATION
Disseminated Intravascular Coagulation (DIC)
Disseminated intravascular coagulation (DIC) is a condition that occurs as a complication of a variety of disorders, including hyperthermia. DIC is characterized by increased intravascular coagulation, worsened by the subsequent formation of microthromboses (clots) that lead to multiorgan failure. The result of DIC is either excessive bleeding or clotting which, without treatment, often leads to death. Animals with DIC typically show signs of bleeding, such as petechiae, ecchymoses, melena, and hematuria. Treatment involves remedying the initial cause of DIC by administering heparin, as well as blood product transfusion to replace consumed coagulation factors. Overall, the treatment or resolution of DIC is many times unsuccessful. DIC has also been called “Death Is Coming.” The best way to prevent DIC is to treat the underlying cause rapidly before the sequence of DIC is initiated.
CLINICAL APPLICATION
Cat Bite Wound
Any animal bite wound has the likelihood of becoming infected. Cat bite wounds, whether on humans or other animals, are especially contaminated and infection is almost guaranteed. The cat’s mouth has a large number of normal flora. Upon penetration of the its teeth through the skin of a person or another animal, those bacteria are deposited deep into the wound where they are no longer normal flora. This deep location often has a low oxygen concentration and becomes an ideal breeding ground for bacteria. The body responds to the presence of these bacteria by sending phagocytic white blood cells to the site to phagocytize the bacteria. This large accumulation of WBCs is known as pus, which often leads to the formation of an abscess (an accumulation of pus in a confined space). Treatment of an abscess typically consists of lancing, draining, and flushing out the wound. Sometimes a drain is placed to allow for further pus to exit the wound. Cat bite wounds, or any penetrating wound, should be addressed quickly to avoid abscess formation. Hint: In the case of bite wounds, look for four puncture sites—one for each of the cat’s canine teeth.
including osmotic pressure changes, edema, and disseminated intravascular coagulation.
PHAGOCYTOSIS. Phagocytosis is one way the body can remove pathogens in the blood, fluids, and body tissues. There are several types of cell capable of phagocytosis but the most common phagocytic cells are the neutrophils, monocytes, macrophages, and dendritic cells. Phagocytes contain receptors on their outer membrane that help differentiate cells as being “self” versus “nonself.” These receptors include PAMP receptors and complement receptors that recognize components of the complement system that enhance phagocytosis.
When tissues are injured or an infection is present, mast cells release histamine, which causes localized vasodilatation and inflammation. Vasodilation creates increased permeability of the capillaries, allowing large numbers of phagocytic white blood cells to enter the injured site.
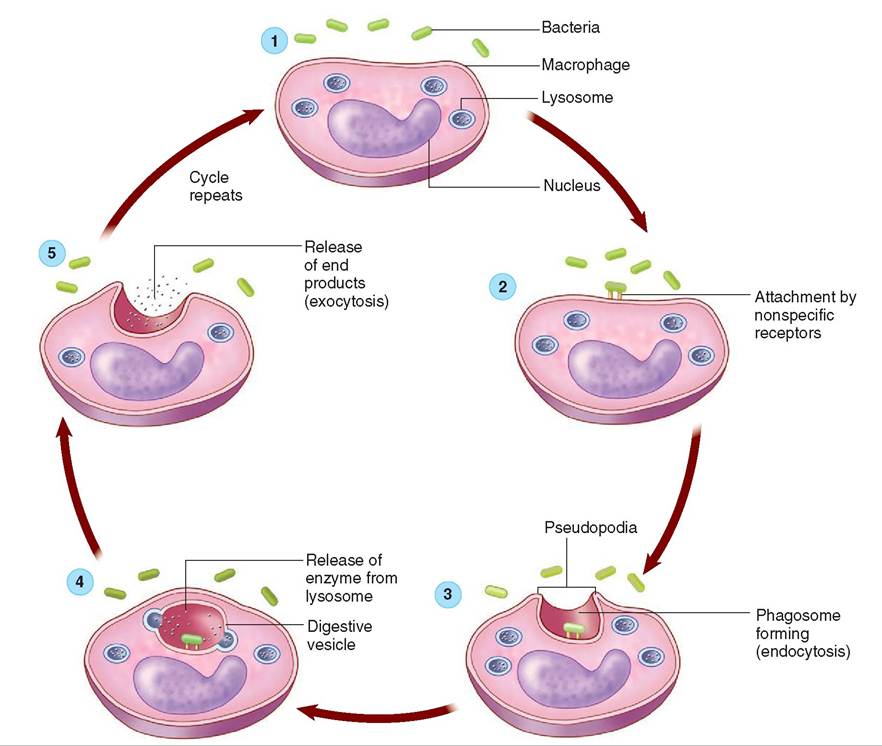
Typically, neutrophils are the first responders to the injured site, followed by macrophages. Phagocytosis includes five steps (Figure 13-9):
1. Activation and chemotaxis: Phagocytes are stimulated by inflammatory signals such as prostaglandins, cytokines, complement proteins, and bacterial components/products to begin moving toward the “nonself” cell (bacterium).
2. Attachment: Receptors on the phagocytes recognize the nonspecific components on the pathogen cell membrane and bind to them.
3. Ingestion or Endocytosis: The attached phagocyte extends projections from its plasma membrane, called pseudopods. Pseudopods engulf the microorganism into a vesicle called a phagosome.
4. Destruction: A lysosome (cellular organelle that contains digestive enzymes to help break down bacteria) fuses with the phagosome, creating a phagolysosome. The lysosome releases digestive enzymes into the phagolysosome. The enzymes break down the bacteria.
5. exocytosis The phagolysosome releases the indigestible material from the phagocyte.
COMPLEMENT SYSTEM. The complement system is a group of 30+ plasma proteins, mostly inactive proteolytic enzymes (break down peptide bonds that hold amino acids together), which are always present in plasma. They become active in the presence of an antigen or an antibody attached to an antigen. They are identified by the letter “C” followed by a number. For example C3 is complement 3. The number indicates the order in which it was discovered. So C3 was the third complement protein discovered.
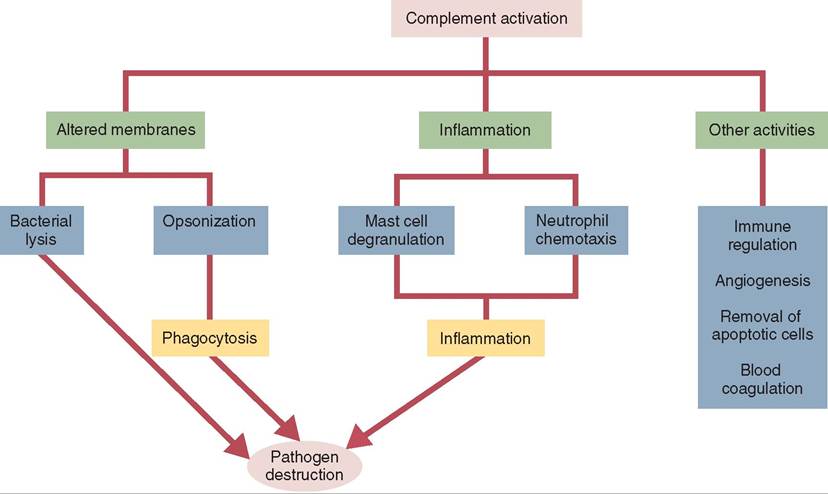
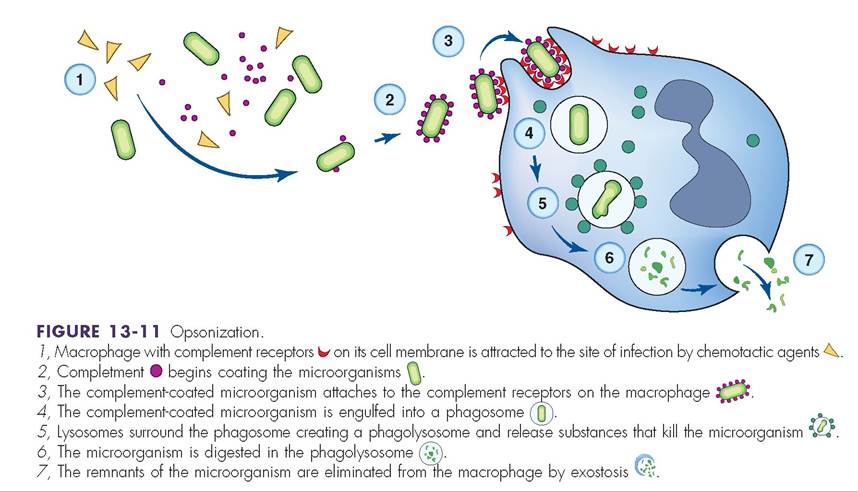
Complement proteins are produced primarily in the liver and circulate in the blood in their inactive form. The two most important functions of the complement system are to trigger inflammation and to alter microbial cell membranes (Figure 13-10). To alter cell membranes the complement proteins can attach to the microbe’s PAMPs and cause cell lysis. These complement proteins are part of the innate immune system because they don’t need an antibody to be bound to the microbe first. The complement response to the PAMPs is rapid and provides nearly immediate protection against the antigen. The complement system can also alter microbe membranes through opsonization (coating the antigen with complement proteins to make it more visible to the phagocyte), for direct destruction by phagocytes (Figure 13-11).
There are also specific complement proteins that require a specific antibody to be bound to a specific antigen so that the specific complement protein can bind to it. This response is slower because it can take from 7 to 10 days for antibody production against an antigen to be effective.
In either case there are a series of complement protein reactions involved in a cascading process whereby when one complement protein is activated it activates the next complement protein in the series. This is called the complement
![]()
FIGURE 13-9 Phagocytosis and destruction of bacteria. (From Patton KT, Thibodeau GA: Anatomy & physiology, ed 8, St Louis, 2013, Mosby.)
![]()
FIGURE 13-10 The functions of the complement system. Complement may either alter microbial membranes or trigger inflammation. Either way, it hastens the elimination of microbial invaders and is thus a key component of the innate immune system. (From Tizard I: Veterinary immunology, ed 9, St Louis, 2013, Saunders.)
![]()
![]()
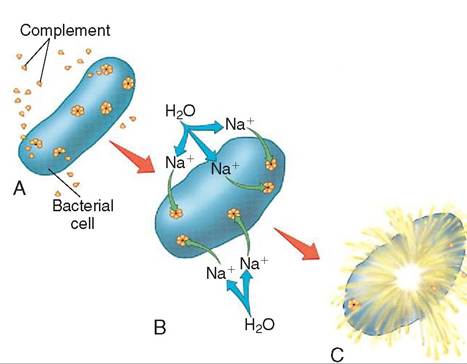
FIGURE 13-12 Complement destruction of a bacterium. A, Activated complement molecules form doughnut-shaped complexes in a bacterium's cell membrane; B, Holes in the complement complexes allow sodium and then water to diffuse into the bacterium; C, After enough water has entered, the swollen bacterium bursts.
cascade. The final step of the complement cascade is complement fixation where the molecules that are formed are gathered in clusters on the antigen's surface. Each cluster resembles a doughnut and in its center a hole is punched in the antigen cell wall. This results in antigen cell lysis or body cell apoptosis (Figure 13-12).
In addition to opsonization, activated complement proteins also play a part in chemotaxis of leukocytes to the site of tissue injury, regulating the inflammatory response and enhancing cell destruction.
CYTOKINES. Cytokines are part of the innate immune system. The term cytokine can be broken down into “cyto” meaning cell and “kinos” meaning movement. Cytokines are communicators. They provide communication between
![]()
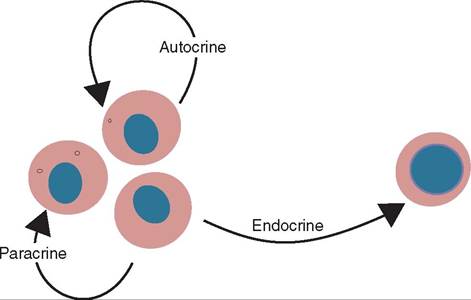
FIGURE 13-13 The distinction among autocrine, paracrine, and endocrine effects. Cytokines differ from hormones in that most of their effects are autocrine or paracrine, whereas hormones usually act on distant cells in an endocrine fashion. (From Tizard I: Veterinary immunology, ed 9, St Louis, 2013, Saunders.)
leukocytes and other cells and among leukocytes themselves. They are signaling proteins that are secreted by certain cells in the body. Cytokines can be autocrine (they act on the cell that secreted them), paracrine (they act on cells near the cell that secreted them), or endocrine (they travel to other parts of the body and act on cells in that location) (Figure 13-13). Often their role is to mediate the immune or inflammatory response by attracting immune cells to a specific site: the site of infection, inflammation, or trauma. They also play a major role in hematopoiesis.
There are many different cytokines but there are groups of cytokines that have similar effects on the cells they target. Some act as inhibitor molecules and others act to enhance immune processes. Many, but not all, cytokines are directly associated with immune activity. Currently nearly 50 cytokines have been described. Interleukins, interferons, and chemokines are types of cytokine.
![]()
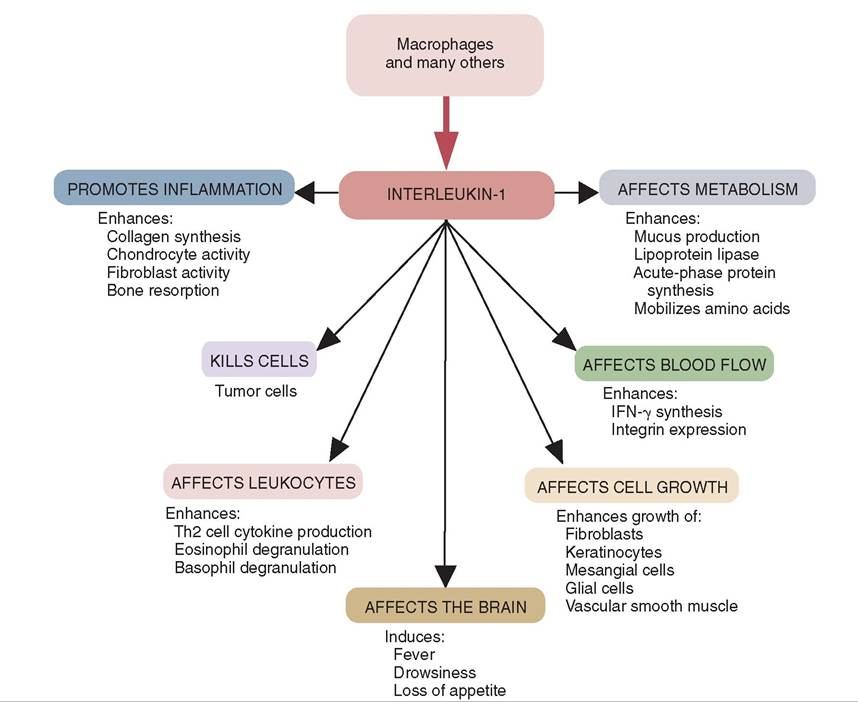
FIGURE 13-14 The origins and some of the biological activites of interleukin (IL)-1. (From Tizard I: Veterinary immunology, ed 9, St Louis, 2013, Saunders.)
INTERLEUKINS. It was initially thought that interleukins were produced by leukocytes and acted on only leukocytes, which is how they got their name. Since that time other cells have been shown to produce interleukins. Interleukins are identified by “IL” followed by a number. Interleukins control leukocyte (especially T and B cells) growth, differentiation, and activation during an immune response. They have a variety of effects in the body. For example, T cells produce IL-2 during an immune response. This interleukin is necessary for the T cells to grow, proliferate, and differentiate into effector (cytotoxic) cells. The activities of IL-1 are shown in Figure 13-14.
CHEMOKINES. Chemokines are chemotactic cytokines. They stimulate the movement of leukocytes from blood into tissue and toward an injury/inflammatory site where there are high concentrations of the chemokines. This often causes an increased rate of clearance of pathogens at an injury site. They have many other effects in the body as well. Many injured or stressed cells will release chemokines, which leads to an influx of immune cells at the site.
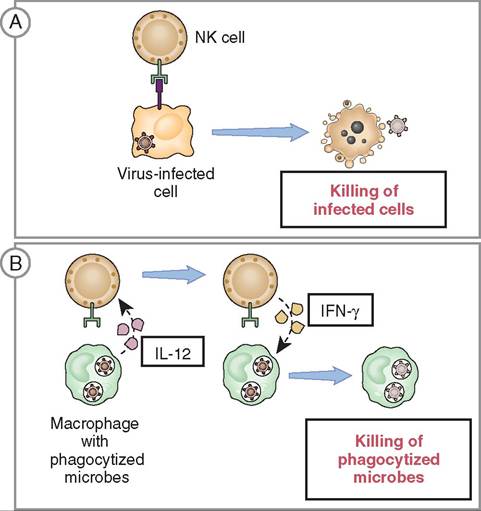
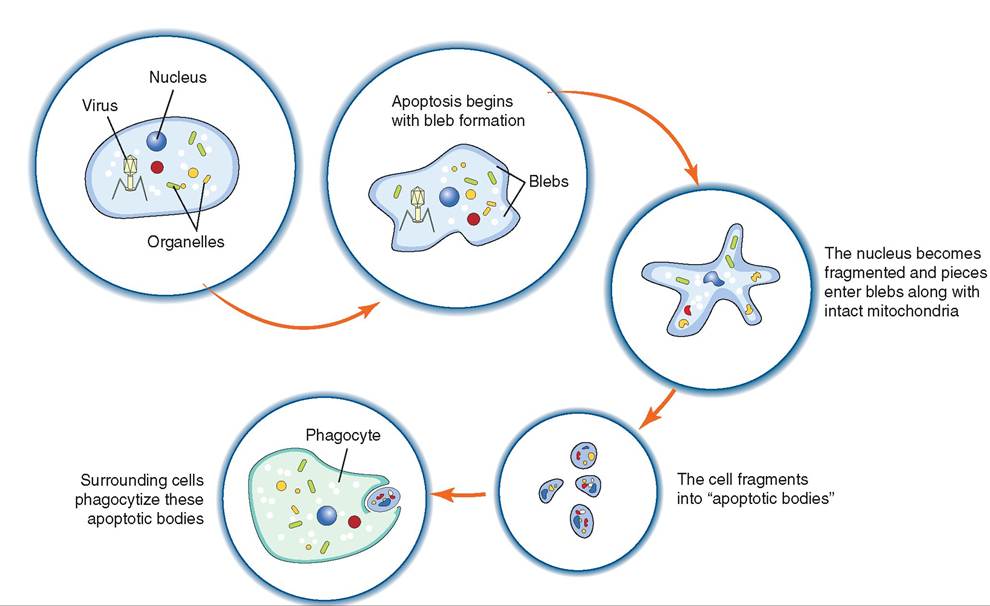
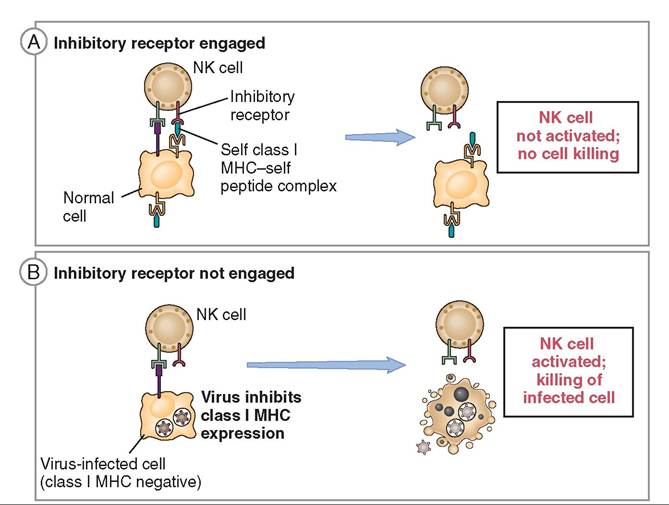
NATURAL KILLER (NK) CELLS. Natural Killer (NK) cells are found in the blood and lymph and are part of both the innate immune system and the adaptive immune system. These granular lymphocytes are able to identify and kill virus- infected cells, stressed cells, and tumor cells (Figure 13-15). They differ from phagocytes in that they do not ingest the target cell. Instead, they bind to the cell and induce cellular changes that lead to apoptosis (programmed cell death) before the virus can mature (Figure 13-16). It is important to note that NK cells do not cause lysis of virus-infected cells, as this would release virions (virus particles) instead of controlling infection. Instead, apoptosis is triggered for virus-infected cells; this ensures death of the virus whereas lysis does not.
All NK cells have two types of receptor on their cell membranes to help determine which cells to kill (abnormal, nonself) or not to kill (normal, self): the killer-activating receptor (KAR) and the killer inhibitory receptor (KIR).
All normal autogenous nucleated cells (self) have MHC-I (major histocompatibility complex class I) molecules on their surfaces that display a normal small protein fragment belonging to the cell (a “self” protein) to which a KIR receptor on an NK cell can bind. This binding demonstrates that the cell is expressing normal amounts of MHC-I and is healthy. This inhibits the NK cell from killing healthy cells (Figure 13-17).
Virus-infected cells or tumor cells in the body often have altered or missing MCH-I. The KIR receptor on the NK cell
![]()
FIGURE 13-15 Functions of natural killer (NK) cells. A, NK cells kill host cells infected by intracellular pathogens, thus eliminating reservoirs of infection. B, NK cells respond to IL-12 produced by macrophages and secrete interferon (INF)-gamma which activates the macrophages to kill phagocytized pathogens. (From Abbas AK: Basic immunology updated edition: functions and disorders of the immune system, ed 3, St Louis, 2010, Saunders.)
cannot bind to an altered or missing MHC-I molecule so it will not inhibit NK cell KAR action. The KAR receptor on the NK cell can still bind with other molecules on the cell membrane of the “nonself” cell, triggering apoptosis.
Once an infected or damaged cell is identified, the KAR eolfl the NK c binds with the cell. The NK cell then releases perforins (proteins that form pores in cell membranes), and proteolytic enzymes such as granzymes from their granules. Granzymes move into the target cell via the newly formed pores and cause apoptosis by breaking down the cell structure and its contents.
NK cells are stimulated by cytokines, including interleukins and interferons. Tumor cells, damaged cells, and virus- ienllfsected c release cytokines as stress signals. NK cells respond, prompting the ultimate destruction of damaged or ienllfse.cted c
INTERFERONS. Interferons (IFNs) are proteins produced by an animal's immune system cells in response to the fpresence o viruses, bacteria, cancer, and other foreign invaders. IFNs are an especially effective mechanism to ward off viral i nvaders. Viruses lack the ability to replicate and survive on their own so they survive by invading the animal's cells ehost cells). While in the host cell, they are able to utilize the ceepllliuclaatrinrg machinery to produce adenosine triphos
phate (ATP) for energy and proteins for viral replication. eVcitriaolnisnf are hard to treat, mainly because they are intracellular pathogens. In order to eradicate the virus, the ehlol st c it inhabits must be damaged.
![]()
FIGURE 13-16 Apoptosis of a virus-infected cell. (From Cunningham, JG: Textbook of veterinary physiology, ed 4, St Louis, 2007, Saunders.)
![]()
FIGURE 13-17 Activating and inhibitory receptors of natural killer (NK) cells. A, Healthy host cells express self MHC-I molecules, which are recognized by inhibitory receptors (KIR), thus ensuring that NK cells do not attack normal host cells. B, NK cells are activated by infected cells when MHC-I is reduced so that the inhibitory receptors are not engaged. The result is that infected cells are killed. (Modified from Abbas AK, Lichtman AH: Basic immunology updated edition: Functions and disorders of the immune system, ed 3, St Louis, 2011, Saunders.)
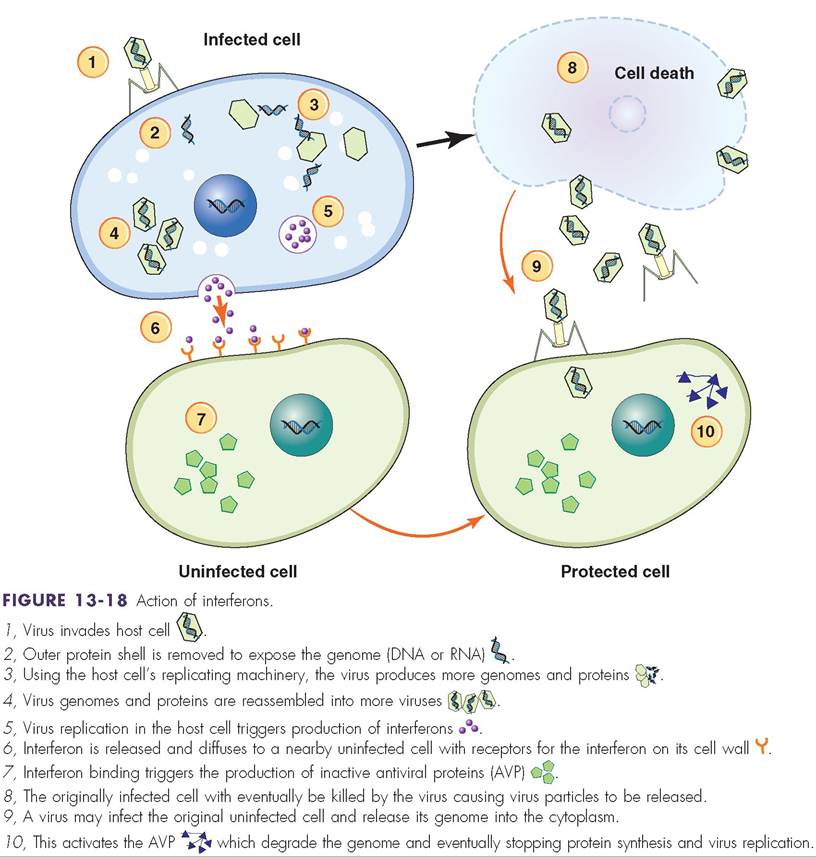
Some virus-infected cells are able to secrete IFNs, which can be beneficial because they prevent the virus from spreading to healthy unaffected cells. Once produced in one cell, IFNs are able to diffuse to neighboring cells and promote the production of “interfering” proteins that help block protein synthesis and degrade viral RNA in the infected cell (Figure 13-18).
More specifically, IFNs are secreted from virus-infected cells and bind to the membrane-bound receptors on surrounding cells. Once the interferon is bound to the noninfected cell, second messengers relay a signal to the inner portion of the uninfected cell to produced inactive antiviral particles (AVP). When a virus enters the cell the inactive AVPs become activated and the overall effect is inhibition of virus replication within that cell.
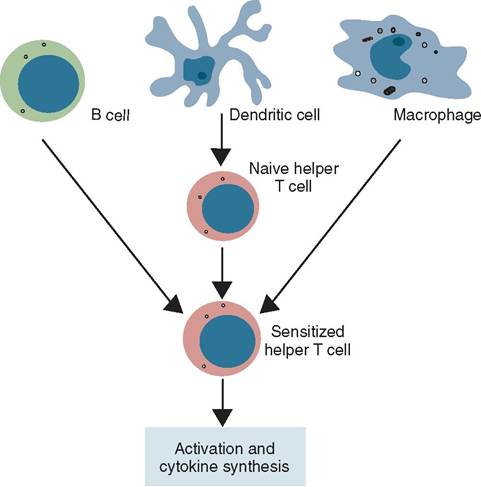
IFNs stimulate an increase in expression of both MHC-I and MHC-II. MHC-II is found on the cell membranes of professional antigen presenting cells (APCs) (Figure 13-19). The APCs are cells whose “profession” it is to phagocytize antigens, process and destroy them, and present fragments of antigen protein attached to an MHC-II on the phagocytic cell membrane. APCs include phagocytes, macrophages, dendritic cells, and B cells. Increased MHC-II expression on antigen-presenting cells leads to recognition by helper T cells to stimulate the action of NK cells and cytotoxic cells.
There are several classes of IFN. Biologically engineered IFNs are now available and have some uses in veterinary medicine, for example in supportive treatment of feline leukemia (FeLV) and feline immunodeficiency virus (FIV).
TEST YOURSELF 13-3
1. What is the difference between the first and second lines of defense against invading pathogens?
2. The body's innate defense against viral pathogens is driven by the production of what?
3. What are the pros and cons of fever?
4. What cell type of innate defense targets tumor cells?
5. What types of cell are phagocytic?
6. What are the four cardinal signs of inflammation?
ADAPTIVE (ACQUIRED) IMMUNE SYSTEM
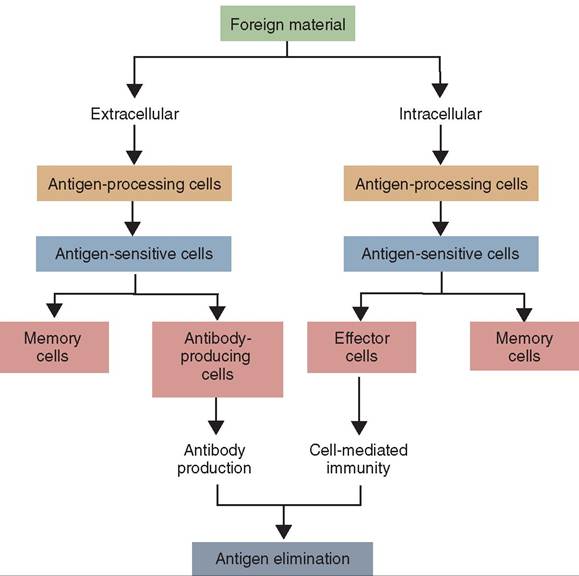
In addition to the nonspecific innate immune system, an animal has a third line of defense that is able to target specific foreign invaders called pathogens or antigens. Unlike the innate immune system, which eliminates anything identified as nonself, the adaptive or acquired immune system is slower to respond, specific, and has memory (Figure 13-20). It targets pathogens with precision. Although the innate immune system protects the body from all pathogens using the same population of cells and chemicals, the cells of the adaptive immune system are programmed to respond only to specific pathogens. The adaptive immune system also has the ability to remember pathogens that have infected the organism and destroy them before they can cause disease in an animal a second time. The adaptive immune system has a systemic, rather than local, impact and is able to respond to pathogens throughout the entire body. Its response to a pathogen involves primarily B lymphocytes and T lymphocytes (Figure 13-21).
![]()
![]()
FIGURE 13-19 Three major populations of antigen-presenting cells (APCs): B cells, dendritic cells, and macrophages. (From Tizard I: Veterinary immunology, ed 9, St Louis, 2013, Saunders.)
![]()
FIGURE 13-20 A simple flow diagram showing the essential features of the adaptive immune responses. (From Tizard I: Veterinary immunology, ed 9, St Louis, 2013, Saunders.)
B LYMPHOCYTES
Like all blood cells, B lymphocytes (B cells) originate in the red bone marrow. They then migrate to lymphoid tissues (e.g., lymph nodes, spleen) where they can initiate an immune response. B cells do not directly destroy pathogens. Each B cell is programmed to secrete a specific antibody (also known as immunoglobulin or Ig) that will lead to phagocytosis and destruction of the pathogen. Since the antibodies that are produced by the B cells are circulating in blood, lymph, and tissue fluid, they are most effective in providing immunity against extracellular pathogens.
(Historical note: The “B” refers to an organ found in birds called the bursa of Fabricius. When B cells were first distinguished from T cells in the 1960s, researchers were working with birds and the bursa of Fabricius was determined to be the site of development. Although mammals do not have this organ, think “B = bone marrow” to remember that B cells mature in the bone marrow.)
B cells are stimulated by the presence of a specific antigen and a signal from a helper T cell to differentiate into plasma cells that are responsible for the actual production, storage, and release of antibodies. Once created, plasma cells remain in the lymph nodes and spleen. Other B cells can differentiate into memory B cells. Should the original pathogen reappear, memory B cells are prepared to respond quickly to the same antigen.
T LYMPHOCYTES
The precursor cells of T lymphocytes (T cells) are thymocytes. These cells originate in the red bone marrow and migrate to the thymus where they mature, multiply, and enter the bloodstream as T cells. They take up residence in the lymph nodes and the spleen, where they coordinate cell- mediated immunity (against intracellular pathogens) and activate B cells.
In an adult animal, B cells and T cells exist in at least three different stages of differentiation:
• Naive cells have entered the lymphatic system, but have not encountered an antigen.
• Cytotoxic or effector cells have been activated and are involved in eliminating a pathogenic antigen.
• Memory cells are the survivors of past infections, capable of providing long-term immunity.
When T cells and B cells are activated in an immune response, they produce clones, which are the memory cells. These clones stay in the lymph nodes or circulate in blood, looking for the same antigen that originally triggered the activation of their parent cell. Should that antigen enter the animal again, memory cells will initiate an immune response. This response is stronger and faster than the initial immune response, hopefully preventing the animal from getting sick.
The adaptive immune system can be divided into two distinct types of immunity, humoral and cell-mediated (Figure 13-22).
HUMORAL IMMUNITY
The humoral immune response, part of adaptive immunity, is triggered by extracellular pathogens and results in antibody production. The antibodies produced by B cells/plasma
![]()
FIGURE 13-21 Development of T and B lymphocytes, and natural killer (NK) cells. (From Tizard I: Veterinary immunology, ed 9, St Louis, 2013, Saunders.)
cells target specific antigens for destruction. Each pathogen has a unique series of amino acid antigen markers on its surface that form a unique shape called an epitope. On the surface of each B lymphocyte there is a corresponding shape that binds to the epitope like two pieces of a jigsaw puzzle. When the B lymphocyte and the antigen bind they wait for a signal from a helper T cell, and then an immune response is initiated. The B lymphocyte creates copies of itself, which will develop into plasma cells that produce an antibody unique to the antigen.
immunoglobins
• IgM antibodies are produced when an animal is first exposed to an antigen (Figure 13-23, A).
• IgM is a temporary antibody that disappears within 2 or 3 weeks after the initial infection and is replaced by the IgG antibody.
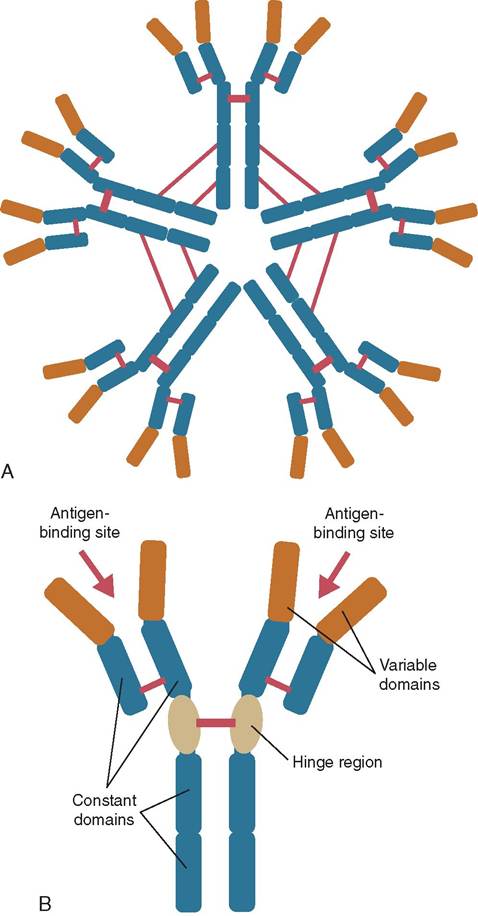
• IgM antibodies are the largest antibody. They are found in blood and lymph fluid and are the first immunoglobulin made by newborn animals.
• IgG is the smallest but most common antibody (Figure 13-23, B). It is produced and released by plasma cells and found in blood and extracellular fluid when the animal has been exposed to an antigen for an extended time.
• Elevated IgG levels can indicate a chronic infection in an animal.
• IgG is the only antibody capable of crossing the placenta to provide passive immunity to the fetus.
• IgG antibodies are involved in fighting bacterial and viral infections.
• Production of IgG antibodies is relatively slow, so the animal may become sick from the initial exposure before the immune response can neutralize the antigen.
• IgA antibodies protect body surfaces from foreign substances. These antibodies play an important role in preventing diseases caused by antigens that may enter the body through mucosal surfaces (e.g., intestinal tract and lungs). Intranasal applications, such as those used in intranasal vaccination, create this type of antibody.
![]()
FIGURE 13-22 Types of adaptive immunity. In humoral immunity, B cells secrete antibodies that eliminate extracellular pathogens. In cell-mediated immunity, T cells either activate machrophages to destroy phagocytosed pathogens or kill infected cells. (From Abbas AK, Lichtman AH: Basic immunology updated edition: Functions and disorders of the immune system, ed 3, St Louis, 2011, Saunders.)
• IgE binds to allergens and triggers histamine release from mast cells and basophils. It also protects against some parasitic helminth (worm) infections.
• IgD leas been shown to activate basophils and mast cells but its exact mechanism is unknown.
B Tpnphocytes and plasma cells remain in lymphoid tissue. The antibodies released by plasma cells are secreted directly into blood, allowing them to find and bind to the invading hntigens. '^^en the antibody binds to the antigen an antigenantibody complex is formed. The results can (1) render antigenic toxins incapable of destroying cells, (2) agglutinate ontigens into large clumps that can be destroyed by macrophages, Ot (3) activate the complement system in plasma tuhrant in t destroys the cell.
CLINICAL APPLICATION
Serologic Testing
Veterinarians often need to distinguish between acute and chronic infecti ons in animals. To help do this, serum samples from a side animal are sent to a reference laboratory where fevels of IgG and IgM are measured. The results help the veterinarian determine whether the illness affecting the eacneinmtal is r or whether it has been around for a long tgiMme. I antibodies are produced early in illness and are usually detectable within 1 to 2 weeks after onset of symptoms. High serum levels of IgM indicate an acute disease. IgG antibodies generally appear 2 to 6 weeks after infection rahunmd hig se levels indicate a more chronic disease.
![]()
FIGURE 13-23 Immunoglobin structure. A, The structure of IgM.
B, The structure of IgG. (From Tizard I: Veterinary immunology, ed 9, St Louis, 2013, Saunders.)
Remember, some B lymphocytes become memory cells after the initial exposure to a pathogen. These cells are capable of protecting the animal from this pathogen should a subsequent exposure occur. In this way, B lymphocytes can both destroy the invading pathogens and create a defense system to protect the body against future infections.
CELL-MEDIATED IMMUNITY
Cell-mediated immunity is also part of the adaptive immune system. It is controlled by T cells, which do not depend on antibody production, but provide immunity against intracellular pathogens. T cells attach directly to antigen markers on the surfaces of phagocytes that have already processed the pathogen.
When T cells are maturing in the thymus, they develop specific antigen receptors on their cell membranes. Each receptor is unique to one specific antigen marker. After being processed in the thymus, T cells travel via the blood to lymph nodes and the spleen. Unlike B cells, T cells leave the lymphoid tissue and circulate throughout the blood and lymph. For this reason, most of the lymphocytes found in peripheral blood are T cells.
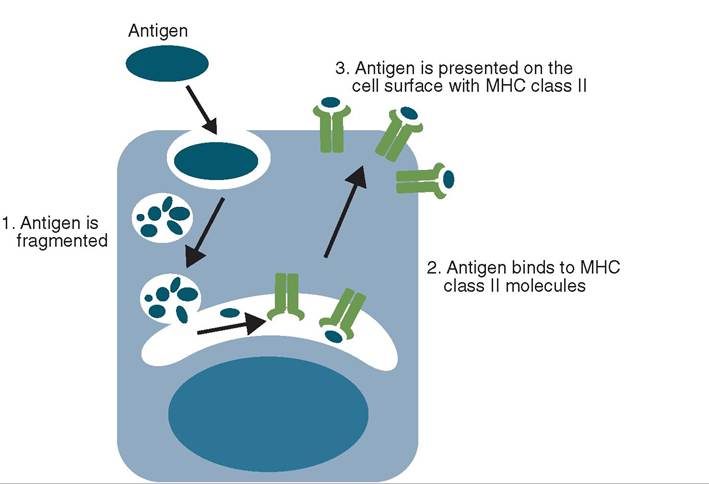
Unlike B cells, T cells fail to recognize antigens on their own. For a T cell to recognize an antigen, it must first be processed by an APC (usually a dendritic cell) (Figure 13-24). The antigen is phagocytized by the APC and broken down to its component parts. Antigenic protein fragments are bound to cellular proteins (MHC-II) in the cell and migrate to the surface of the APC. The T cell recognizes the combination of MHC-II and the antigenic marker on the macrophage surface as the antigen to which it was programmed in the thymus during development.
When the T cell encounters a corresponding antigenic marker on an APC, the antigenic marker and T cell bind, resulting in the T cell becoming activated, or sensitized. The sensitized T cell then divides, creating two populations of new cells. The first population contains multiple clones of the first cell, called memory T cells. The second population subdivides into three distinct populations:
• Helper T cells (Th): These are the most numerous T cells.
They help the immune response by secreting cytokines into the surrounding tissue. The effects of these signaling proteins include:
• The cytokine that increases activation of B cells, cytotoxic T cells, and suppressor T cells
• Interleukins, such as interleukin 2 (IL-2), a cytokine that stimulates the activity of other T cells
• Macrophage migration inhibiting factor (MIF), a cytokine that attracts tissue macrophages to the affected area via chemotaxis. This activates the macrophages to accelerate the rate of phagocytosis that will result in more antigenic markers on the surface of the macrophage. MIF also inhibits the antigens from leaving the site of infection.
• Cytotoxic T cells (Tc): These cells are also known as effector cells, killer cells, or killer T cells. They attach to antigenic markers and destroy the cells to which they are attached. However, they are not damaged themselves. T cells that do not become cytotoxic cells can become helper T cells.
• Regulatory T cells (Ts): These cells inhibit helper T cell and cytotoxic T cell function by negative feedback. They also prevent B cells from transforming into plasma cells. These antagonistic actions provide a certain degree of control over the cell-mediated and humoral immune responses.
ACTIVE IMMUNITY
Active immunity is, as the name implies, the result of an active immune process. During an initial exposure to an antigen, the immune response can be slow, allowing the disease to develop. During subsequent exposures to the same antigen, the immune system recognizes the antigen and can respond faster. Using memory T and B cells, the body
![]()
FIGURE 1 3-24 The processing of exogenous antigen by an antigen-presenting cell. (From Tizard I: Veterinary immunology, ed 9, St Louis, 2013, Saunders.)
produces antibodies, protecting the individual from developing the disease a second time.
In natae, animals are exposed to a variety of antigens in their environment and develop a natural immunity against these antigens without ever showing signs of disease. In a similar manner, using vaccines we can protect animals aergtaaiinnst c diseases without ever exposing the animals to tchtueaal disease.
There are two primary types of vaccine used in veterinary medicine today: modified live and killed virus. Modified live vaccines contain like-virus particles that have been weakened (attenuated) so they are nonpathogenic but still recognized by the animal as antigenic. These inactivated particles are unlikely to cause disease. A modified live vaccine produces a sutnroeng imm response because they mimic a natural infection.
Killed virus vaccines contain virus particles that have rbeeaetnedt with chemicals, heat, or radiation to render them inactive. Once killed, the viruses are unable to revert to their disease-causing state, but the immune system is still eaocbolegntizer the remnants of the virus and respond
appropriately. This response is weaker than when modified laicvceinves are used so multiple doses must be administered before immunity is achieved. Regardless of whether exposure is through the environment or vaccination, the body's immune system actively responds to the antigens, resulting in active immunity.
PASSIVE IMMUNITY
Receiving antibodies from an external source can also protect the animal but its immune system is not actively involved. This is passive immunity. The most common method of providing passive immunity is through maternal antibodies
CLINICAL APPLICATION
Vaccination Protocols 1
Occasionally a client will come into a veterinary clinic and ask to hwe a pet vaccinated so it can go to the groomer, boarding facility, or daycare facility. Or a client wants some livestock vaccinated so it can be shipped across a border that same day. H is important for clients to understand that although vaccines will protect their animals from disease, the protection is not immediate. Vaccines stimulate the immune system (active immunity), but it takes time, usually about 2 weeks. As veterinary technicians, we need to help educate clients so they can help keep the animals in their care healthy aond up t date on vaccines.
tehat ar passed from mother to the fetus through the placenta (tssnsplscentally), or to the neonate via colostrum (the antibody-rich milk the mother produces right before and after toh ). Although these antibodies will protect the newborn animal, they do not activate the immune system amnodryme cells are not produced. Once the antibodies against a specific disease are cleared from the newborn's system it is no longer protected from the disease.
MECHANISMS OF DISEASE
Iyn a health animal, once exposure to a pathogen has occurred, the innate immune system should immediately recognize the pathogen as nonself and begin its attack. Inflammation, phagocytosis, and fever should work together to destroy the pathogen, without regard to its type. Bacterial, viral, and fungal infections elicit similar initial responses from the innate immune system.
CLINICAL APPLICATION
Vaccination Protocols 2
Vaccination protocols in young animals are established using the characteristics of both passive and active immunity. The newborn animal likely has some passive immunity from its mother (transplacentalIy or through colostrum) that will protect it against commonly encountered antigens. As the newborn matures the passively received antibodies are lost raontde cption must be activated by vaccinations and the aucnteive imm system. There is no way to know when the passive immunity disappears so initial vaccinations usually require a smes of injections.
For example, most puppies receive their initial vaccination ser^^^ιere around 4 to 6 weeks of age. Prior to this time we assume the puppy is protected by passive immunity. When it is about 6 weeks old the puppy must start providing its oi∙eeι antibodies through active immunity. To make sure rewoet eacrtinpg the puppy as best we can we will continue
providing vaccines up to about 16 weeks of age.
Most vaccines do not provide lifetime immunity so the animal’s immune system must be “boosted” at regular intervals, e.g. annually, every 2 years, or every 3 years.
TEST YOURSELF 13-4
1. Where αreB cells produced? Wheredothey mature?
2. Where ared cellsprodeced?Wheredo they mature?
3. Whichcell prodoieeantibodies?
4. Describe the three stages of differentiation of lymphocytes.
5. Describe the function of each of the five immunoglobulins. When would the levels of each increase?
6. Describe the three types of T lymphocyte.
7. Describe how vaccines protect patients from disease. Is this an example of active or passive immunity?
The adaptive immune system also responds to the pathogen. Although its response is slightly delayed, it is specific to the invading pathogen and is able to target it more directly. Typically, this multifaceted defense system is sufficient to prevent an animal from becoming seriously ill. However, there are cases where the pathogen is too strong or the immune system is too weak and the animal succumbs to disease.
The following factors determine the likelihood of a pathogen causing disease in the animal:
• Exposure: The process of a pathogen infecting a host begins with exposure to the pathogen. In order to invade a host, the animal must be exposed to the pathogen. Exposure can occur via a variety of methods including contact with contaminated secretions (aerosolized or nonaerosolized), direct ingestion of the pathogen, or contact via a break in the skin (wound). In all of these rceases, the has been a failure of the first and second lines of deeeιιse that are designed to prevent the pathogen from gnatrinying e into the body.
• Mode of infection/transmission: l-hwh^^^ts that use
raaenrosmsoilstsion to move from one animal to another have an easier time finding and invading the bodies of potential hosts. If, however, transmission requires direct contact with another infected animal, transmission is less likely and the pathogen may have a harder time infecting subsequent hosts.
• Virulence: The virulence of the pathogen and the degree of pathogenicity refer to the relative strength of the pathogen. Some pathogens are weak and are easily destroyed by the immune system. Others are able to resist trhoetepctive methods of the immune system and cause illness in the animal.
• Immune system strength: The immune systems of young, old, or immunosuppressed animals are usually not functioning at 100% capability. Also, animals fighting a concurrent disease are more susceptible to disease. If an animal’s immune system is using its resources to fight one pathogen, there are fewer resources available to fight the second pathogen. This increases the chance that the second pathogen will cause disease.
• Resistance: There are various types of resistance that can
hniemlpaal a remain protected from a particular pathogen. Acquired resistance is the most common. It is acquired onvimeraal’s a lifetime, either through natural exposure tahotehtogepn or via vaccination against the pathogen. In abhsoeetsh, c t immune system has been primed, through emllemory c production, to fight the pathogen, making subsequent encounters less likely to result in disease. Some eavneimr als n develop a disease when exposed to a particu-
laatrhopgen, even if they have never been vaccinated. These animals have resistance based on their DNA rather than ⅛eir ^ime exposure. Species resistance protests all members ot' a species from some diseases. For example, dogs do not contract human measles and people do not get sick after exposure to canine parvovirus.
HYPERSENSITIVITY REACTIONS
The immune system is critical to an animal’s survival, but occasionally it malfunctions, resulting in mild health disturbances, major illnesses, or even death. What we recognize as an Mlengic reaction is actually an overreaction of the immune system. Instead of the usual controlled response, the immune system goes into overdrive. This results in the signs we commonly associate with allergies.
Innsistiezed animals, allergic reactions can become even vmeorer,e se resulting in hypersensitivity reactions, which
laarsesicfied into four types.
• Type I reactions nre generally severe and can include
anaphylactic shock (urticaria or hives, edema, excess sali- voamtiiotin,gv, dyspnea, diarrhea, cyanosis, shock, and
deayh). Atopy (atopic dermatitis), flea allergy dermatitis, and food allergies in dogs and cats are examples of type Ienarctions. I type I reactions, antigens bind to the surface igobEfoIdiesant on the surfaces of basophils and mast cells rather than IgA, IgG, or IgM. This sensitizes the animal. Once sensitized, a second exposure to the same easnutiltgsen r in the release of inflammatory chemicals lionotodstthreabm, which leads to the symptoms often associated with severe allergic reactions and anaphylactic shock. Without immediate medical intervention, these reactions can quickly become fatal.
• Type II reactions occur ⅛n infection is present, antibodies are being produced, and the complement system is activated. Cross-reactive antibodies form, leading to antibody-mediated cytotoxic reactions (AMCR). This rcecaucrtsion o when reactive antibodies bind antigens on the host cell surface and destroy the body's own cells. doTshis lea t diseases where the immune system is the fcause o disease rather than the cure. These reactions vooftlevne in red blood cells, leading to diseases such as immune mediated hemolytic anemia (IMHA) where the animal's immune system destroys its own red blood cells indiscriminately, causing a life-threatening anemia.
• Type III reactions occur ⅛n an antibody and an antigen boirnmd and f an immune complex. These immune com- eplexes ar insoluble and become trapped in the basement membrane of small blood vessels in tissues, especially joints, kidneys, lungs, brain, and skin. Activation of the complement cascade leads to chemotaxis of a large number of neutrophils and other inflammatory cells and chemicals to the site. This typically produces acute inflammation and damage to affected tissues. Type III reactions are some of the most common immunologic diseases.
Examples include systemic lupus erythematosus, hyper- sneenusmitiovnitiytisp, and vasculitis.
• Type IV reactions. Type IV or cell-mediated reactions occur ⅛n antigens trigger helper T cells (Th), which in tcutirvnatea cytokines, macrophages, and cytotoxic T cyeplels. T IV reactions are often caused by intracellular pathogens and are accompanied by inflammation. An fexample o a Type IV reaction is systemic inflammatory response syndrome (SIRS). SIRS is a form of shock characterized by an exaggerated inflammatory response, usually ⅛e to a severe infection or extensive tissue damage. Athough most commonly seen during bacterial infections, other pathogens (such as fungi, viruses, and parasites) can all cause SIRS.
TEST YOURSELF 13-5
1. Whiat threefactors inflUencesthes abilityofa pathogen to caarc dircarc?
2. Describe ths three typcr of rcrirtancc that prevent an animal from contracting a disease. Which type of resistance prevents hamanr from contracting canine distemper?
3. Which hyperseasitivity reactienio Iiedly i naolved in a vaccine reaction?
∕j CLINICAL APPLICATION
Immune Mediated Hemolytic Anemia
Lucy, a 23 lb, 7-year-old female spayed Cocker Spaniel, presented with acute lethargy/weakness, anorexia, exercise intolerance, and hematuria. According to her owner, Lucy was vaccinated 3 weeks prior to the onset of clinical signs. Lucy appeared depressed in the exam room and was quiet, but alert aesnpdornsive.
Based on the diagnostic test results the veterinarian diag- uosed Lucy with immune mediated hemolytic anemia (IMHA) and autoimmune thrombocytopenia (ITP), likely secondary to vcctmction. Lucy was hospitalized and placed on intravenous (IV) fluids, glucocorticoids (to prevent destruction of lreoellodsdb c and suppress the overactive immune system), iavnedn was g a transfusion of whole blood to treat anemia.
iPtahtients w IMHA and thrombocytopenia may have coagulopathies, which will interfere will blood clotting, so a large vessel like the jugular vein should not be used to draw blood samples. Caution stickers were also placed on Lucy's cage stating “NO JUGULAR BLOOD SAMPLES.”
What Are IMHA and ITP?
Immune mediated hemolytic anemia is a immune malfunction where the body destroys its own red blood cells (RBCs), either W creating antibodies directed at its own RBCs, or by IgG and complement binding to the RBCs marking them for destruction.
eThere ar two types of IMHA, primary and secondary. Primary IMHA occurs when the body creates antibodies directed at its own RBCs. Secondary IMHA occurs when foreign proteins bind to RBC membranes (e.g., FeLV in cats, vaccines, Ehrlichia canis infection, ⅛ns, zinc toxicity, onion troxicity, o heartworm infection).
A diagnosis of IMHA is based on a combination of clinical signs and diagnostic test results. Patients typically present with lethargy, weakness, pale mucous membranes, icterus, hematuria, bruisme/petechiae, and possibly vomiting and diarrhea. Examination often shows hepatosplenomegaly, due to removal oC increased numbers of destroyed RBCs from circulation.
Icterus is caused by the excess destruction of RBCs, which releases mo bilirubin into the bloodstream than the liver can choisnjugate. T results in the bilirubin being deposited in skin auncodums membranes and giving them a yellow tinge.
Blood tests will show hyperglobulinemia (excessive antibod- ihnreecsuilatiton, c indicating an active immune system),
hyperbilirubinemia, anemia, reticulocytosis (increase reticulo- bgcolor=white>INTRODUCTION, 339
THE HEART, 340
Location, 340
Size and Shape, 340
Coverings of the Heart, 340
Wall of the Heart, 341
Chambers of the Heart, 341
Valves of the Heart, 343
Skeleton of the Heart, 343
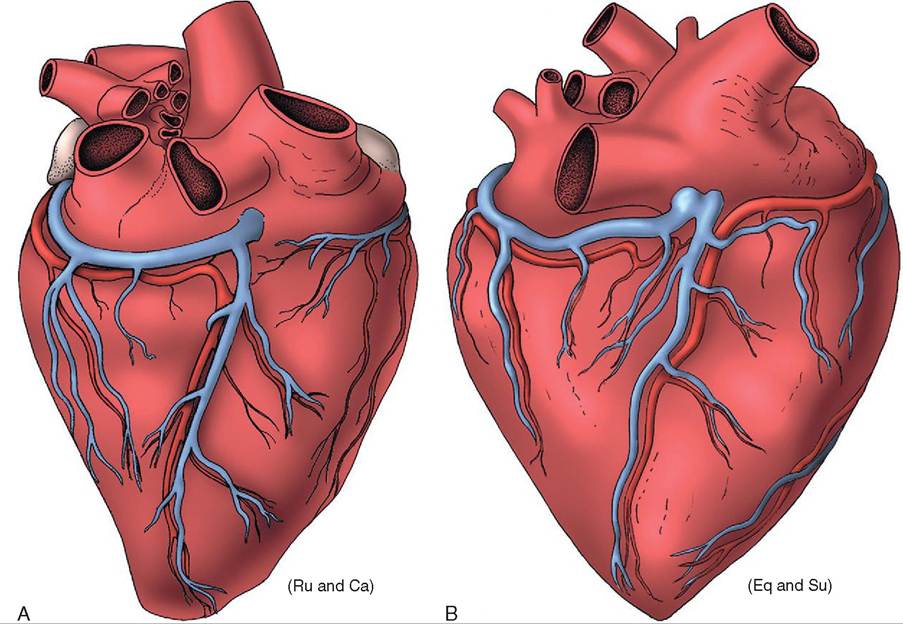
Blood Supply to the Heart, 344
Nerve Supply to the Heart, 344
Blood Flow Through the Heart, 345
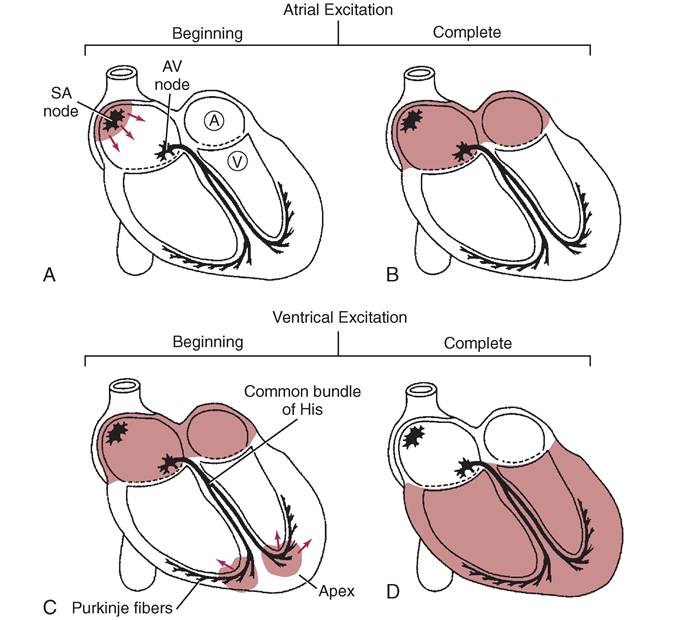
CARDIAC CONDUCTION SYSTEM, 348
NORMAL HEART SOUNDS, 350
ABNORMAL HEART SOUNDS, 351 CARDIAC OUTPUT, 351 BLOOD VESSELS, 352 Arteries, 352 Capillaries, 353 Veins, 353 BLOOD CIRCULATION IN THE FETUS, 354 PULSE, 357 Pulse Points, 357 BLOOD PRESSURE, 357 CARDIOVASCULAR MONITORING, 358 Electrocardiography, 358 Echocardiography, 358 VENIPUNCTURE, 359 | | LEARNING OBJECTIVES | |
When you have completed this chapter you will be able to:
1. Describe the external and internal anatomy of the heart.
2. Follow the flow of blood through the heart, pulmonary circulation, and systemic circulation.
3. Explain how the heart valves keep blood flowing in the proper direction through the heart.
4. Describe the components of the cardiac conduction system and explain how it works to keep the heart beating in an organized fashion.
5. Explain what happens during one cardiac cycle.
6. Understand cardiac output and what conditions can affect it.
7. Describe the anatomy of arteries, veins, and capillaries and understand the function of each type of blood vessel.
8. Understand the difference between fetal and newborn circulation.
9. Understand the different methods used to evaluate the cardiovascular system.
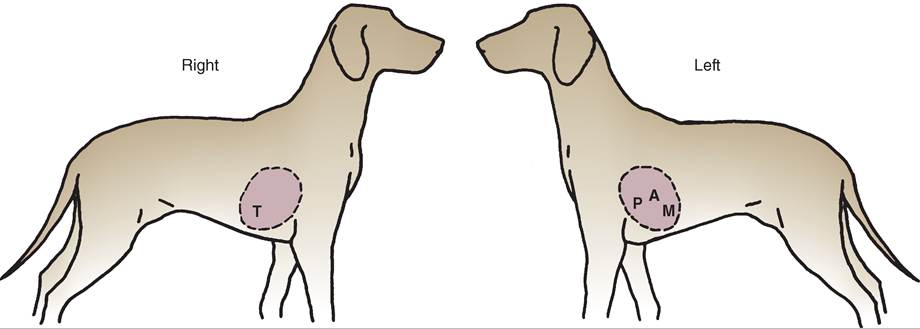
10. Know the common pulse points and venipuncture sites for common species of animal.
VOCABULARY FUNDAMENTALS
Afterload ahf-tar-lod
Aorta a-ohr-tah
Aortic valve a-ohr-tihck vahlv
Apex of heart a-pehcks of hahrt
Arteriole ahr-teer-e-ol
Artery ahr-tar-e
Atrioventricular node a-tre-o-vehn-trihck-u-lahr nod
Atrioventricular septum a-tre-o-vehn-trihck-u-lahr sehp-tuhm
Atrioventricular valve a-tre-o-vehn-trihck-u-lahr vahlv
Atrium a-tre-uhm
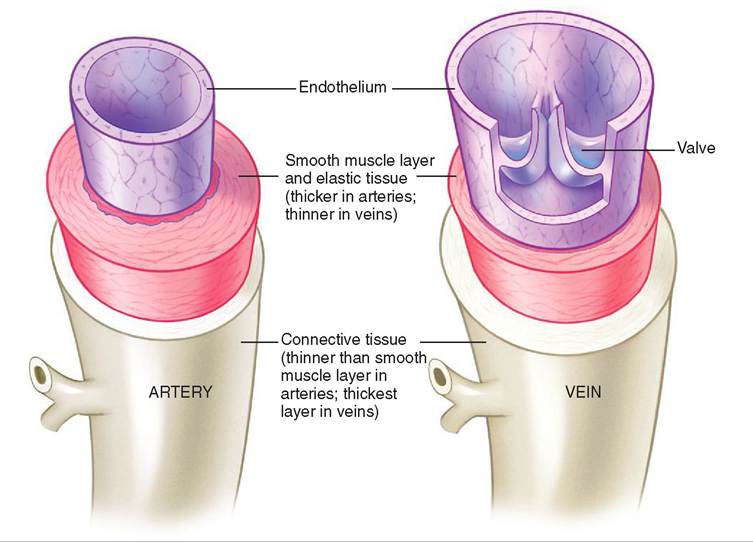
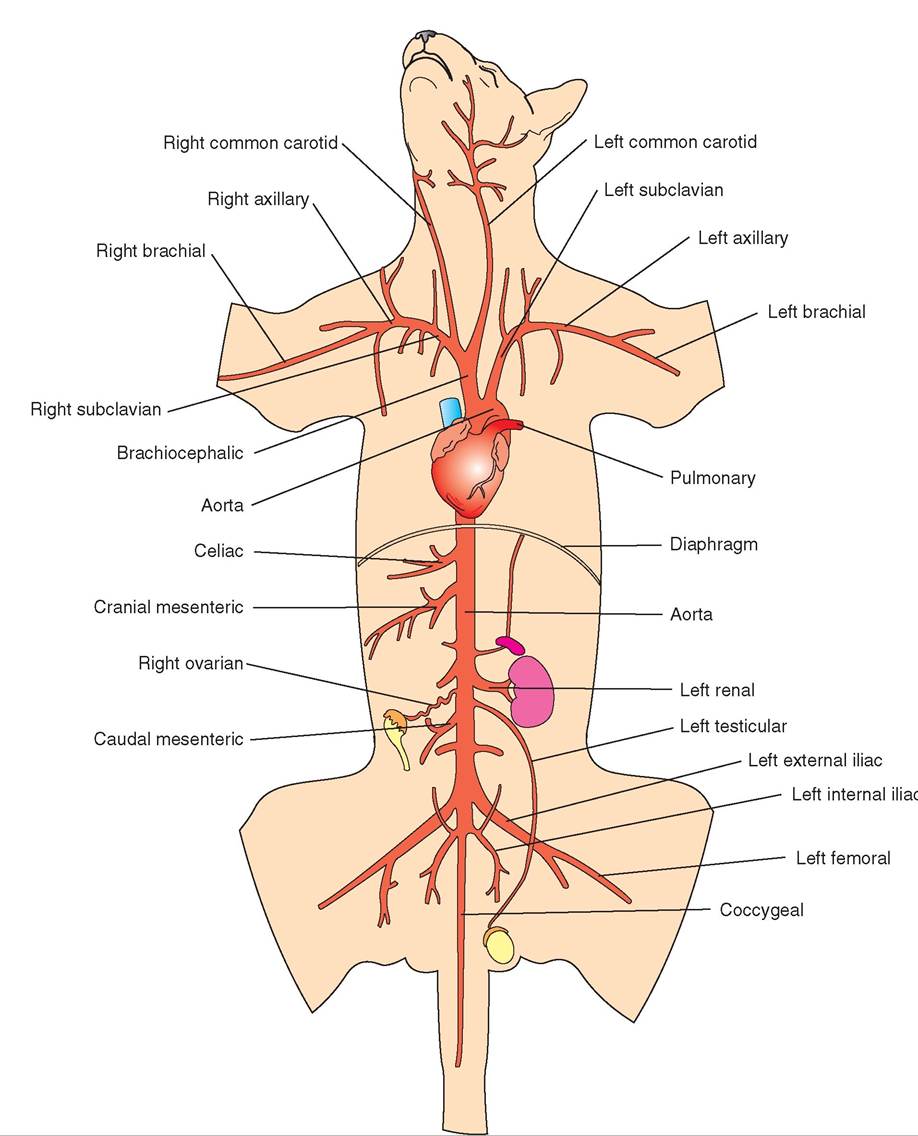
Auricle ohr-eh-kuhl
Auscultation aws-kuhl-ta-shuhn
Autorhythmic aw-to-rihth-mihck
Base of heart bas of hahrt
Bicuspid valve bι-kuhs-pihd vahlv
Blood pressure bluhd prehsh-ar
Bundle of His buhn-duhl of hihs
Capillary kahp-eh-lahr-e
Cardiac cycle kahr-de-ahck sι-kuhl
Cardiac output kahr-de-ahck out-puht
Cardiovascular system kahr-de-o-vahsk-u-lahr sihs-tehm Carotid artery kahr-oht-ihd ahr-tar-e
Cephalic vein seh-fahl-ihck van
Chordae tendonae kohr-da tehn-duhn-a
Coccygeal vein kohck-sehj-e-ahl van
Coronary artery kohr-ah-nar-e ahr-tar-e
Coronary sinus kohr-ah-nar-e sι-nuhs
Coronary vein kohr-ah-nar-e van
Cusp kuhsp
Deoxygenated de-ohck-seh-jeh-na-tehd
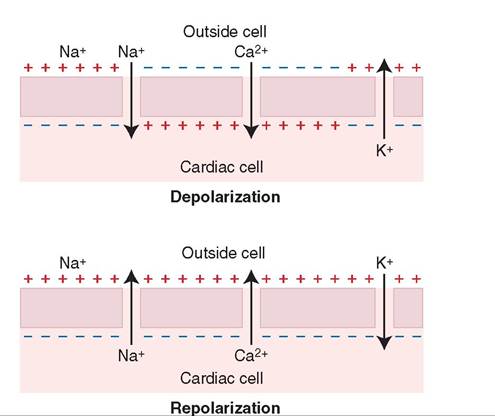
Depolarization de-pδ-lar-ih-za-shuhn
Diastole dι-ahs-stδ-le
Diastolic blood pressure dι-ah-stohl-ihck bIuhd prehsh-ar
Doppler ecaocardiography dohp-lar ehck-δ-kahr-de-ohg-rah-fe
Ductus arteriosus duhck-tuhs ahr-teer-e-δ-suhs
ECHO ehck-δ
EcCocardiograpCy ehck-δ-kahr-de-ohg-rah-fe
Elastic artery eh-lahs-tihck ahr-tar-e
Electrocardiogram e -lehck-trδ-kahr-de-δ-grahm
ElectrocardiograpCy e-lehck-trδ-kahr-de-ohg-rah-fe
Endocardium ehn-dδ-kahr-de-uhm
EndotCelium ehn-dδ-the-le-uhm
Epicardium ehp-ih-kahr-de-uhm
Femoral vein fehm-ohr-ahl van
Foramen ovu/e fohr-a-mehn δ-vah-le
EIeart rate hahrt rat
Interatrial septum ihn-tar-a-tre-uhl sehp-tuhm
Interventricular groove ihn-tar-vehn-trihck-u-lar grvov
Interventricular septum ihn-tar-vehn-trihck-u-lar sehp-tuhm
Jugular vein juhg-u-lar van
Me∣an arterial pressure men ahr-teer-e-ahl prehsh-ar
Mediastinum me-de-ah-stιn-uhm
Mitral valve mι-trah vahlv
Murmur mar-mar
Muscular artery muhs-kyoo-lar ahr-tar-e
Myocardium mι-δ-kahr-de-uhm
Oscillometric aws-uh-lδ-meh-trihck
Oxygenated ohck-suh-jehn-a-tehd
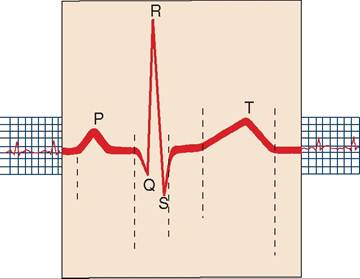
P wave P wav
Papillary muscle pah-pihl-lear-e muhs-uhl
Parietal layer of tCe serous pericardium pah-rι-eh-tahl la-ar of the seer-uhs pear-ih-kahr-de-uhm
Pericardial fluid pear-ih-kahr-de-ahl floo-ihd
Pericardial sac pear-ih-kahr-de-kahl sahc
Pericardial space pear-ih-kahr-de-ahl spas
Pericardium pear-ih-kahr-de-uhm
Polarization pδl-ar-ih-za-shuhn
Preload pre-lδd
Pulmonary artery puhl-muh-near-e ahr-tar-e
Pulmonary circulation puhl-muh-near-e sar-kyoo-la-shuhn
Pulmonary valve puhl-muh-near-e vaviv
Pulse puhls
Pulse rnle puhls wav
Purkinje fiber system par-kihn-je fl-bar
sihs-tehm
QRS complex Q-R-S kohm-plehkx
Repolarize re-po-lar-ιz
SapCenous vein sahf-uh-nuhs van
Semilunar valve seh-me-lu-nar vahlv
Serous pericardium see^uhs pear-ih-kahr-de-uhm
Sinoatrial node sι-nδ-a-tre-ahl nδd
SpCygmomanometer sfihg-mδ-muh-nohm-uht-ar Starling’s law stahr-lihngz lahw
Stroke volume strδk vohl-um
Systemic circulation sihs-tehm-ihck sar-kyoo-la-shuhn
Systole sihs-tuh-le
Systolic blood pressure sih-stohl-ihck bluhd prehsh-ar
Systolic discharge sih-stohl-ihck dihs-chahrj
T wave T wav
Tricuspid valve trι-kuhsapihd valv
Umbilical artery uhm-bihl-ihck-ahl ahr-tar-e
Umbilical vein uhm-bihl-ihck-ahl van
Valvular Insufficiency vahl-vyoo-lar ihn-suh-fihsh-ehn-se
Valvular stenosis vahl-vyoo-lar stelι-no-sihs
Vein van
Vena cava ve-nah ka-vah
Ventricle vehn-trihck-ehl
Venule vehn-yool
Virceral layer of tCe serous pericardium vih-sar-ahl la-ar of the seer-uhs pear-ih-kahr-de-uhm
INTRODUCTION
Imagine the world blood lives in. Think of it as a water world. The plasma is the fluid all the elements live and swim in. The outer limits of this world are the walls of the blood vessels where blood resides. The erythrocytes (red blood cells) are the planes, trains, and automobiles that move oxygen and other substances from place to place. The leukocytes (white blood cells) are the military vehicles ready for battle at a moment's notice. The thrombocytes (platelets) are the EMTs, the first responders to the scene of a vessel wall injury.
So far this is a static world; nothing is moving. Enter the heart. Each time the heart beats blood is propelled through blood vessels throughout the animal's body. This is the world of the cardiovascular system (i.e., the circulatory system). It is responsible for the movement of blood and everything it carries throughout the animal's body. It is made up of the heart, all the blood vessels, and the blood itself. Normally there are no external openings to the cardiovascular system so it is considered a closed system. Electrolytes, waste materials, nutrients, hormones, antibodies, and drugs are carried by blood contained in the structures of the cardiovascular system to every living cell in the animal's body.
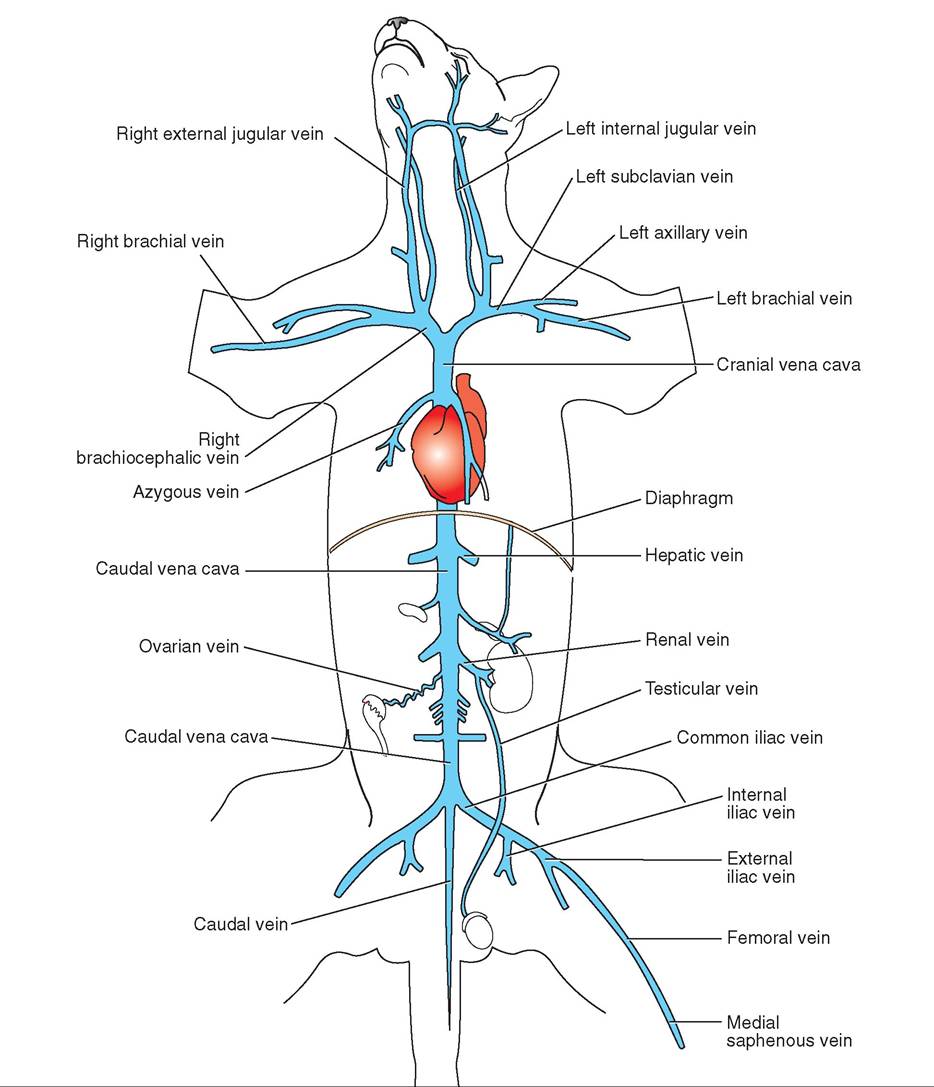
Blood is continuously flowing around the animal's body and through the heart in a circuit propelled by the beating (pumping) heart. Arteries carry blood away from the heart; veins carry blood toward the heart; and capillaries form the transition between arteries and veins.
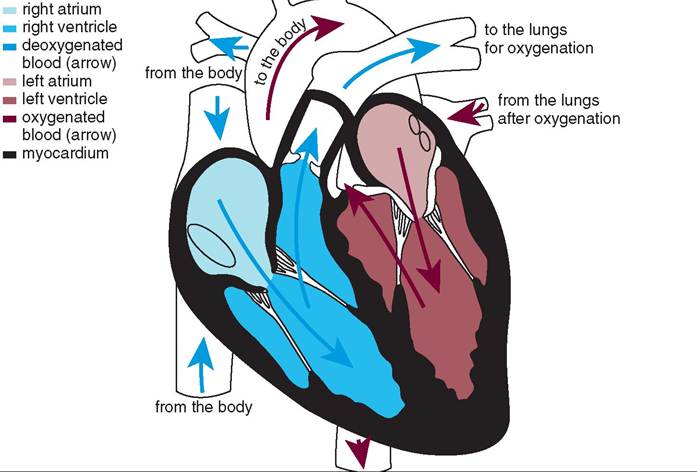
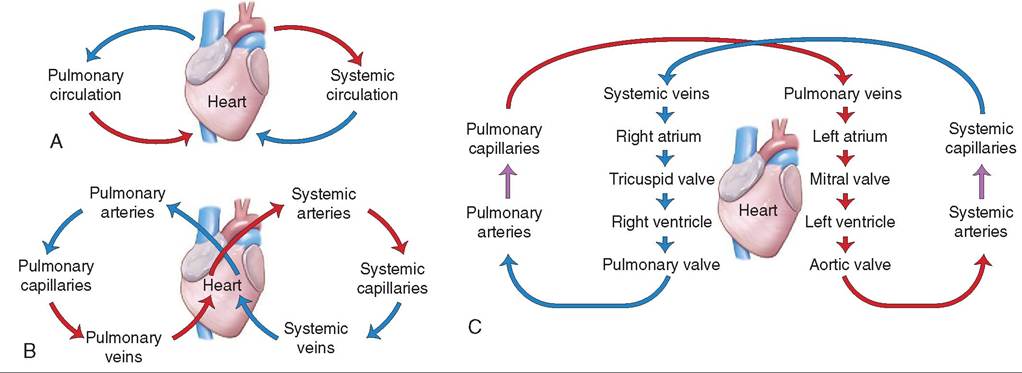
The cardiovascular system is divided into two parts that all blood cycles through in a “figure 8” configuration: the pulmonary (lung) circulation and the systemic (body) circulation. One side of the heart controls each part. The right side of the heart controls the pulmonary circulation. It receives deoxygenated blood from throughout the animal's body (carried in veins) and pumps it into the lungs where it becomes oxygenated. The left side of the heart controls the systemic circulation. It receives oxygenated blood from the lungs and pumps it out to the rest of the animal's body. More on this later.
THE HEART
LOCATION
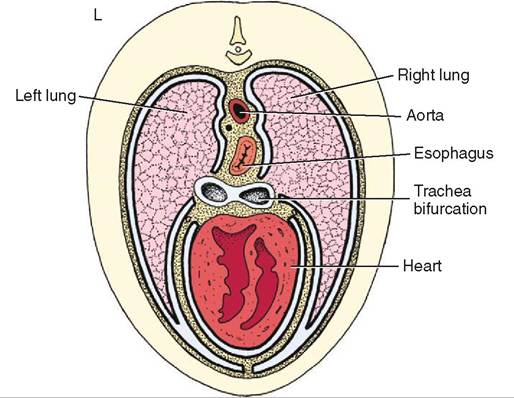
The heart is located in the middle of the thoracic cavity in the mediastinum, the space between the two lungs (Figure 14-1). The mediastinum in bounded by the thoracic inlet crainially, the diaphragm caudally, the sternum ventrally, and the spinal column dorsally. In addition to the heart, the mediastinum also contains blood vessels, the thoracic portion of the trachea, the esophagus, the thymus in young animals, lymph nodes, and nerves.
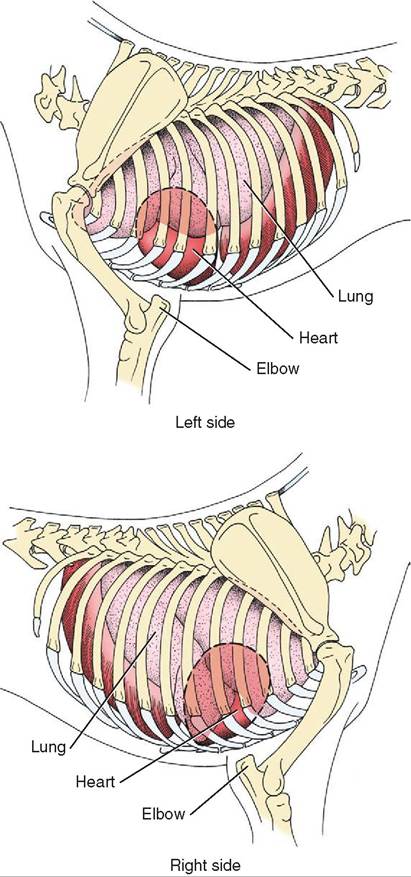
Generally speaking, when viewing a standing animal the heart is located between the elbows (Figure 14-2).
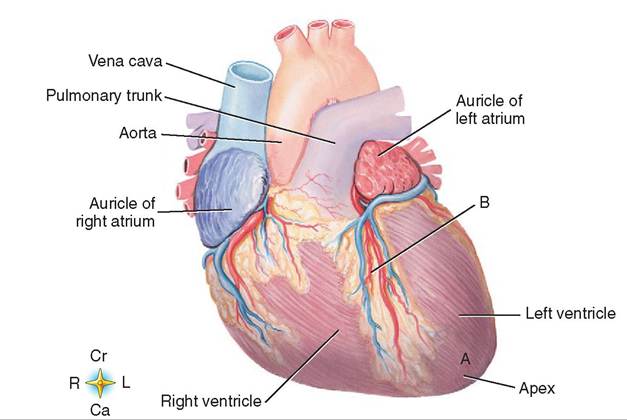
SIZE AND SHAPE
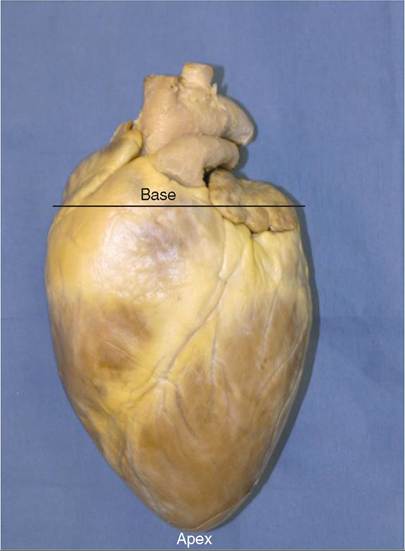
The heart is sort of heart-shaped (Figure 14-3). It has a rounded cranial end called the base of the heart. The more pointed caudal end is the apex of the heart. This is just the opposite of what we normally think of base (wide bottom) and apex (narrow top) but if you go strictly by shape and forget orientation the wide end is the base and the narrow end is the apex.
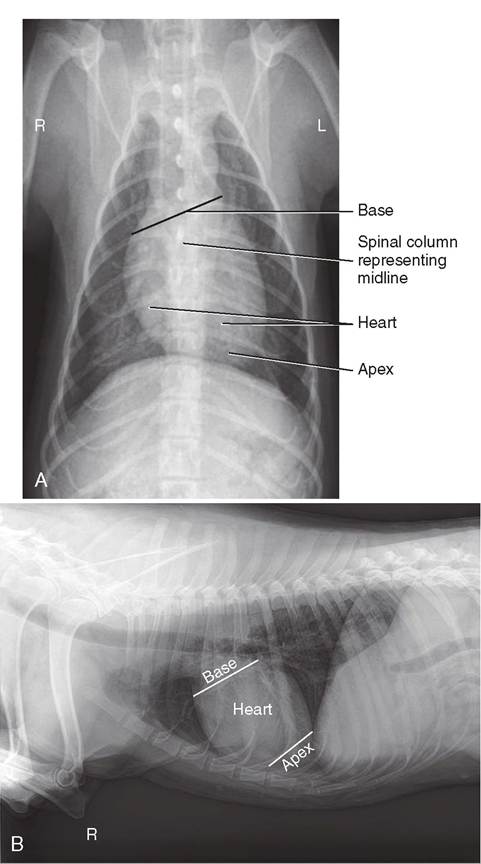
The heart doesn't sit straight along the median plane in an animal. The base is shifted to the right and faces more dorsally. The apex is shifted to the left and sits more ventrally (Figure 14-4).
![]()
FIGURE 14-1 Transverse section through the thorax at the level of the heart showing structures in the mediastinum. (Redrawn from Dyce KM, Sack WO, Wenseng CJG: Textbook of veterinary anatomy, ed 4, St Louis, 2010, Saunders.)
COVERINGS OF THE HEART
The heart is contained in a fibrous sac called the pericardium. The pericardium is divided into two parts: the fibrous sac called the pericardial sac and the serous pericardium. The pericardial sac is a little loose so the heart can beat inside it but it is not elastic so it cannot stretch if the heart becomes abnormally enlarged.
![]()
FIGURE 1 4-2 Location of the heart between the elbows in a standing animal. (Redrawn from Dyce KM, Sack WO, Wenseng CJG: Textbook of veterinary anatomy, ed 4, St Louis, 2010, Saunders.)
![]()
FIGURE 14-3 Shape of the heart showing the base and the apex.
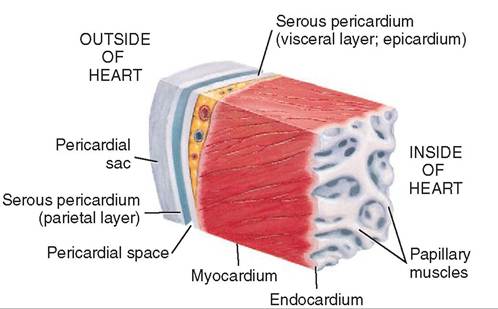
The serous pericardium consists of two membranes. A smooth, moist serous membrane called the parietal layer of the serous pericardium lines the pericardial sac, and the visceral layer of the serous pericardium lies directly on the surface of the heart. The pericardial space is the area between the two serous membranes. It is filled with pericardial fluid that lubricates the two membranes and prevents friction as they rub together during contractions and relaxations of the heart muscle (Figure 14-5).
WALL OF THE HEART
The wall of the heart has three layers (Figure 14-6). The middle and thickest layer is the muscular layer called the myocardium because it is made up of cardiac muscle. Remember that cardiac muscle fibers are joined side-to-side by multiple branches and end-to-end by intercalated discs. These two anatomic characteristics mean that the myocardium is made up of continuous muscle sheets that wrap around the chambers of the heart. These muscle sheets make a greater force of contraction possible.
Two other advantageous characteristics of cardiac muscle are that it is autorhythmic and it doesn't fatigue. This means that without outside stimulus it can start beating (contracting and relaxing) in a steady rhythm before an animal is born and continue beating through birth, adolescence, adulthood, middle age, and old age without taking a break. When the heart stops beating and it isn't restarted the animal dies.
The epicardium is the outermost layer of the heart wall. It is a membrane that lies on the external surface of the myocardium. Sound familiar? Another name for
![]()
FIGURE 14-4 Radiograph showing the position of the heart in the thoracic cavity. A, ventral view; B, lateral view. (A, From Evans H, Lahunta A: Miller's Anatomy of the Dog, ed 4, St Louis, Saunders. B, From Brown M, Brown: Lavin's Radiography for Veterinary Technicians, ed 5, St Louis, 2014, Saunders.)
the epicardium is the visceral layer of the serous pericardium. Two names; same membrane.
The endocardium is the membrane that lies on the internal surface of the myocardium. It is composed of thin, flat simple squamous epithelium and forms the lining of the heart chambers. The endocardium is continuous with the endothelium that lines blood vessels. The endocardium also covers the valves that separate the chambers of the heart.
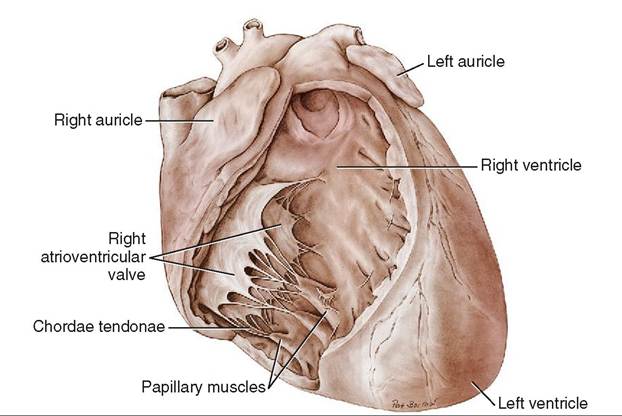
The inside surface of the myocardium is not smooth. It forms ridges and nipplelike projections called papillary muscles that are covered by the endocardium.
CHAMBERS OF THE HEART
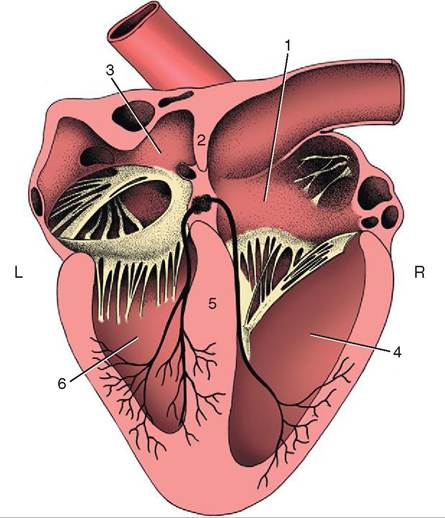
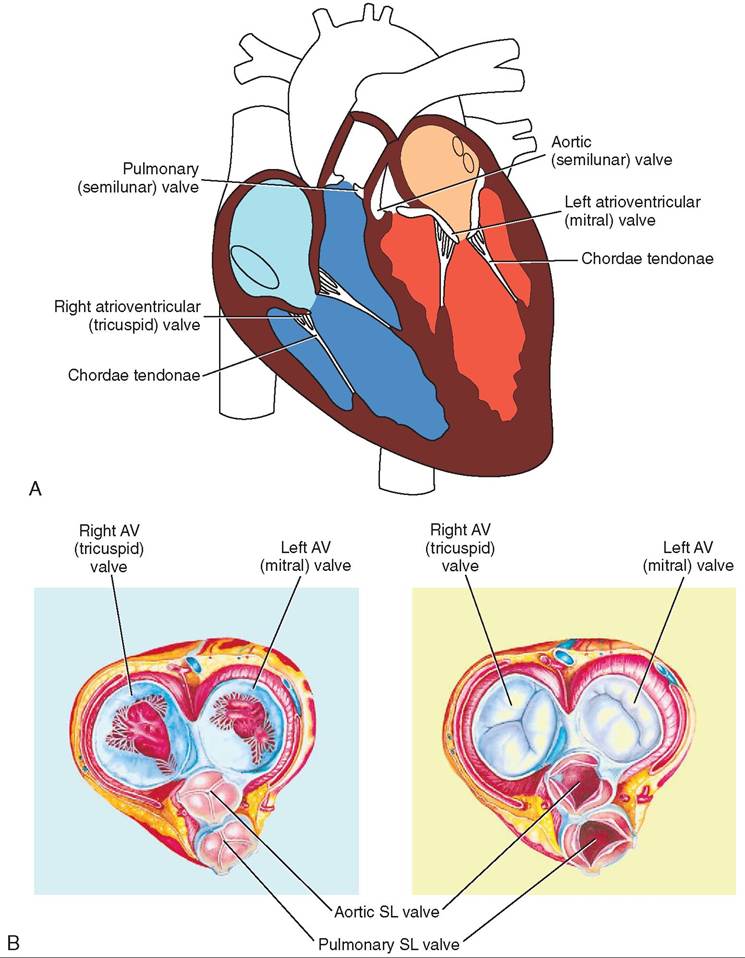
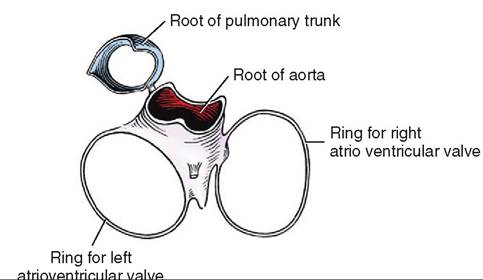
There are four chambers or cavities in the heart: two atria (singular: atrium) that receive blood into the heart, and two ventricles that pump blood out of the heart (Figure 14-7). The
![]()
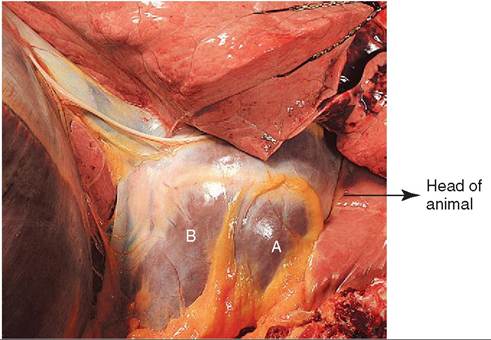
FIGURE 1 4-5 The heart enclosed in the pericardium. A, The wall of the right ventricle visible through the pericardium; B, the wall of the left ventricle visible through the pericardium. The heart is sitting in the area of the mediastinum. (Modified from Clayton HM, Flood P, Rosenstein D: Clinical anatomy of the horse, London 2005, Mosby Ltd.)
![]()
FIGURE 14-6 Section of the wall of the heart showing the pericardial sac, the parietal and visceral layers of the serous pericardium, the pericardial space, the myocardium, and the endocardium. (Modified from Huether SE, McCance KL: Understanding pathophysiology, ed 4, St Louis, 2007, Mosby.)
TEST YOURSELF 14-1
1. Which type of blood vessel carries blood away from the heart? Toward the heart?
2. What are the two parts of the cardiovascular system? Which part carries blood to and from the left rear leg of a pony?
3. List three structures found in the mediastinum.
4. Which is located more caudally in a standing pig, the apex or the base of the heart?
5. What is the difference between the endocardium and the pericardium?
two atria sit on top of the two ventricles and their walls form part of the base of the heart. The two ventricles sit below the two atria and the wall of the left ventricle forms the apex of the heart.
ATRIA
The left atrium and the right atrium are separated by the interatrial septum that is a continuation of the myocardium. The atria receive blood from large veins that carry
![]()
FIGURE 14-7 Section of the heart exposing the four chambers. 1, Right atrium; 2, interatrial septum; 3, left atrium; 4, right ventricle; 5, interventricular septum; 6, left ventricle. (Modified from Dyce KM, Sack WO, Wenseng CJG: Textbook of veterinary anatomy, ed 4, St Louis, 2010, Saunders.)
blood to the heart. When the atria have filled with blood their walls (composed of myocardium) contract and force blood through one-way valves into the ventricles.
The atria are identified on the outside of the heart by their auricles. These are blind pouches that come off the main part of the atria and look like earflaps.
That’s Interesting: Auricle means “ear flap” or “ear.” In humans the auricle also refers to the external ear flap. Since the atrial auricle looks somewhat like an earflap it seems to make sense to name it accordingly.
The auricle is part of the atrium but is not the entire atrium so the two terms cannot be used interchangeably.
The myocardium of an atrium is not very thick because it only has to contract with enough force to move blood into a ventricle. The same is not true for the walls of the ventricles.
VENTRICLES
The left and right ventricles are separated by the interventricular septum, which is a continuation of the interatrial septum. Together they form the atrioventricular septum. The area of the interventricular septum is visible on the outside of the heart as the interventricular groove (Fi^ιre 14-8). The groove contains coronary (heart) blood vessels and is frequently filled with fat. When the ventricles have received blood from the atria the myocardium of the ventricular walls contract and force blood through one-way valves into arteries. The right ventricle pumps blood to the pulmonary circulation through the pulmonary artery; the left ventricle pumps blood into the systemic circulation through the aorta. Since blood from the right ventricle doesn't have very far to go the right ventricular wall is thinner than the left ventricular wall. The left ventricular wall has the most work to do pumping blood to the rest of the animal's body so it has a thicker wall that will contract with greater force. In fact the left ventricular wall is so thick that it pushes the right ventricular wall to the right. Consequently the left ventricular wall makes up the apex of the heart (Figure 14-9).
VALVES OF THE HEART
There are four one-way valves that control blood flow through the Iieart (Figure 14-10). Two of the valves are located between the right and left atria and their respective ventricles. The other two are located between the right and left ventricles and the arteries they eject blood into. The valves close at specific times to prevent backflow of blood into the chamber it just
![]()
FIGURE 14-8 Left view of the heart showing the interventricular groove filled with fat. (Modified from Clayton HM, Flood P, Rosenstein D: Clinical anatomy of the horse, London 2005, Mosby Ltd.)
came from. In order for the heart to function properly blood must flow through it in one direction only.
The atrioventricular valves (AV valves) are located between the atria and the ventricles. The right AV valve consists of three flaps or cusps of endothelium and is called the tricuspid valve. It opens when the pressure from the amount of blood in the right atrium forces it open and allows blood to flow into the right ventricle. When the pressure from the blood in the right ventricle exceeds the pressure of blood in the right atrium the tricuspid valve is forced to snap shut. The valve is prevented from opening backward into the atrium by collagen fiber cords that are attached to the edge of each cusp and to papillary muscles in the wall of the right ventricle. These cords are called the chordae tendonae LFignre 14-11).
The left AV valve has only two cusps and is called the bicuspid valve. It is also known as the mitral valve because of its resemblance to the headgear, called the miter, worn by Roman Catholic bishops. This valve also has attached chordae tendonae.
The two valves that control blood flow out of the ventricles and into arteries are the semilunar valves, so named because they have three cusps, each of which resembles a crescent moon. The right semilunar valve is the pulmonary valve because blood from the right ventricle flows through it into the pulmonary circulation. The left semilunar valve is the aortic valve because blood from the left ventricle flows through it into the aorta, which is the major artery that is the beginning of systemic circulation.
SKELETON OF THE HEART
The skeleton of the heart is located between the atria and the ventricles (Figure 14-12). It is made up of four dense fibrous connective tissue rings and has four primary functions:
• It separates the atria and ventricles.
• It anchors the heart valves.
• It provides a point of attachment for the myocardium.
• It provides some electrical insulation between the atria and the ventricles.
![]()
FIGURE 14-9 Ventral view of the heart showing A, the wall of the left ventricle forming the apex of the heart and B, the interventricular groove. (Modified from Brown M, Brown L: Lavin's Radiography for veterinary technicians, ed 5, St Louis, 2014, Saunders.)
![]()
FIGURE 14-10 Valves in the heart. A, Longitudinal slice through the heart. B, Cross section of the heart. AV, Atrioventricular; SL, semilunar. (B, From Thibodeau D: Structure and function of the body, ed 14, St Louis, 2012, Mosby.)
BLOOD SUPPLY TO THE HEART
Like any living tissue, the cells of the heart need nourishment and oxygen brought to them and waste materials carried away. Coronary arteries and coronary veins are responsible for these functions (Figure 14-13). Coronary arteries branch off the aorta just past the aortic valve (left semilunar valve). They continue to branch around the heart until they completely encircle it. The left ventricle gets the largest blood supply because it has the most work to do pumping blood through the systemic circulation. After the blood from the coronary arteries has passed through the capillaries in the myocardium it enters the coronary veins. To return the blood to the circulation the coronary veins join together near the right atrium and form a channel called the coronary sinus that drains directly into the right atrium.
NERVE SUPPLY TO THE HEART
Cardiac muscle is autorythmic and can create its own contractions and relaxations through its internal conduction system. But there are times when the heart needs to beat faster to increase the oxygen supply to certain tissues. For example, an animal being exercised needs an increased oxygen supply to its muscles. To help accommodate this increased oxygen demand the heart receives some external
![]()
FIGURE 14-12 Skeleton of the heart. (Modified from Evans H, Lahunta A: Miller's anatomy of the dog, ed 4, St Louis, 2013, Saunders.)
![]()
FIGURE 14-11 Interior view of the right ventricle showing the chordae tendonae of the right AV (tricuspid) valve. Only two cusps of the valve are visible. (Modified from Evans H, Lahunta A: Miller's anatomy of the dog, ed 4, St Louis, 2013, Saunders.)
motor stimulation. The nerve fibers enter the heart and terminate primarily in the right atrium near the area of cardiac muscle cells that is the control center for the cardiac conduction system. Some nerve fibers will stimulate an increased heart rate and others will stimulate a decreased heart rate. The nervous system is discussed in detail in Chapter 9. The nerve supply to the heart serves a purpose, but is not essential. Transplanted hearts lose their nerve supply and usually continue to function well.
TEST YOURSELF 14-2
1. Which sits closer to the base of the heart, the left atrium or the right ventricle?
2. What is the name of the structure that is a continuation of the myocardium that forms a wall between the two atria? The two ventricles?
3. Why is the wall of the right ventricle thinner than the wall of the left ventricle?
4. What is another name for each of these valves: right AV valve, semilunar valve in the right ventricle, left AV valve, semilunar valve in the left ventricle?
5. What is the function of the chordae tendonae?
CLINICAL APPLICATION
Hardware Disease
In cattle, the reticulum (the most cranial stomach compartment) rests directly behind the heart, and the two organs are separated by the muscular diaphragm. Cattle are not very selective when eating, and it is not uncommon for them to ingest wires, nails, and other foreign metallic objects along with their feed. These bits of “hardware” are ingested into the rumen, from which digestive contractions move them forward into the reticulum. Continued ruminal contractions, particularly when combined with factors that increase abdominal pressure, such as pregnancy and parturition, may push pieces of wire through the cranial wall of the reticulum. Puncture of the reticulum wall by a foreign object often results in traumatic reticuloperitonitis, also called hardware disease, which is an inflammation and infection of the reticulum and abdominal cavity. More severe disease occurs when a wire is pushed even farther cranially, through the diaphragm and into the pericardium. This can result in septic pericarditis, which is an infection of the pericardium that usually progresses to heart failure and death.
Hardware disease can be prevented by the oral administration of a magnet about the size of a 5 ml blood tube. The magnet stays in the rumen or reticulum, usually for the rest of the animal’s life. Wire or other metal objects ingested by the animal stick to the magnet instead of being pushed crani- ally through the wall of the reticulum and beyond.
BLOOD FLOW THROUGH THE HEART
You’ve read about heart chambers, one-way valves, and unidirectional blood flow so now it’s time to follow the blood as it moves through the heart. The entire purpose of the heart is to receive deoxygenated blood from the systemic circulation (right atrium), send it out to the pulmonary circulation for oxygenation (right ventricle), receive the freshly oxygenated blood back from the pulmonary
![]()
FIGURE 14-13 Coronary circulation of the heart viewed from the right. A, Ruminants and carnivores, B, horse and pig. Ru, Ruminants; Ca, cat; Eq, equine; Su, pig. Red blood vessels, coronary arteries; blue blood vessels, coronary veins. (Modified from Dyce KM, Sack WO, Wenseng CJG: Textbook of veterinary anatomy, ed 4, St Louis, 2010, Saunders.)
CLINICAL APPLICATION
Pericardial Effusion and Cardiac Tamponade
The heart is able to expand and contract in the chest thanks to the layer of fluid that provides lubrication between layers of the serous pericardium. Normally, only a small amount of fluid is contained within the pericardial sac. A number of conditions such as infection, inflammation, or hemorrhage may cause excess fluid to accumulate in the pericardial sac. This condition is called pericardial effusion. Sometimes pericardial effusion is idiopathic, meaning it may occur spontaneously with no known cause.
The outer layer of the heart, called the fibrous pericardium, is not elastic, so when the pericardial space is overfilled with fluid, the heart becomes unable to expand normally between contractions. This condition, called cardiac tamponade, leads to less complete cardiac filling, decreased stroke volume, and decreased cardiac output. Pericardial effusion, with or without cardiac tamponade, may be treated by inserting a needle into the pericardial sac (usually through the chest wall) and withdrawing the excess fluid.
circulation (left atrium), and put it into systemic circulation (left ventricle) (Figure 14-14). Note that each chamber has its own specific function. Think of blood flow as a figure 8. One loop represents the pulmonary circulation, the other loop represents the systemic circulation, and the heart sits in the middle pumping blood through both loops (Figure 14-15).
We'll arbitrarily start our journey through the heart in the vena cava, the large vein that brings deoxygenated blood from the systemic circulation (but not the pulmonary circulation) to the heart. The vena cava enters the right atrium of the heart. The deoxygenated blood passes from the right atrium through the tricuspid valve into the right ventricle. At this point the pulmonary valve in the right ventricle is closed. When the right ventricle is full the tricuspid valve is forced closed and the pulmonary valve is forced open. The right ventricle contracts and the deoxygenated blood leaves the right ventricle through the pulmonary valve and enters the pulmonary circulation via the pulmonary artery. After the deoxygenated blood has been oxygenated in the pulmonary circulation it comes back to the heart via the pulmonary vein that empties into the left atrium. The mitral valve opens and the blood from the left atrium enters the left ventricle. When the ventricle is full the mitral valve is forced closed and the aortic valve is forced open. The left ventricle contracts and oxygenated blood leaves through the aortic valve and enters the systemic circulation via the aorta, the largest artery in the body. Once in the systemic circulation the blood is distributed throughout the animal's body through arteries that become progressively smaller until they transition to become capillaries. At the tissue capillary level, blood gives up its oxygen in exchange for carbon dioxide and
![]()
FIGURE 14-14 Card iac blood flow. Arrows indicate the direction of blood flow. (From Colvile J, Oien S: Clinical veterinary language, St Louis, 2014, Mosby.)
![]()
FIGURE 14-1 5 Schematics of the circulation. A, Pulmonary and systemic circulation, B, vessels of the pulmonary and systemic circulation, C, blood flow through the heart plus the pulmonary and systemic circulation.
other waste materials in tissue fluid. From this tissue level the capi^ies converge to form small veins that become progressively larger as the deoxygenated blood is carried back to the heart. The veins eventually join the vena cava and the deoxygenated blood flows into the right atrium. The coymclpeleistec.
euToremak s blood is entering and leaving the heart in an orde^l^, coordinated fashion, the heart has to have rhythmic, ∞ordinated contractions. The actual rhythm pattern is simple. fen though the two sides of the heart pump blood to different areas they do so at the same time: deoxygenated bnltoerosd e the right atrium and oxygenated blood enters the left atrium at the same time; the tricuspid and mitral
valves open and close at the same time; the right and left ventricles contract at the same time, and so forth. When the eventricles ar full the pressure from the blood in them forces the AV valves closed and the semilunar valves open. When the pressure in the pulmonary artery (right side) and aorta (left side) exceeds the pressure in the right and left ventricles tmhiel usne ar valves are forced close. At this point the pres
sure in the atria exceeds the pressure in the ventricles so the AV valves open and ventricular filling begins again. To keep bwlionogd, flo the ventricles are emptying while the atria are filling and the atria are emptying while the ventricles are hfielling. T valves closing produce sounds that can be heard with a stethoscope: the heartbeat.
![]()
TEST YOURSELF 14-3
1. List, in order, the structures an erythrocyte will pass through (including valves) to move in a complete circuit from the left ventricle back to the left ventricle.
CLINICAL APPLICATION
Defibrillation
If you’ve watched a few hospital shows on television, chances are you’ve seen someone grab a couple of paddles, yell “clear” or something like it, place the paddles on the patient’s chest, and “shock” them to restart the heart. In real life, this process is called defibrillation, and it has to do with the electrical conduction system of the heart.
Sometimes a diseased heart will develop one or more ectopic pacemakers. The word ectopic means out of place, and an ectopic pacemaker is located outside the heart’s normal pacemaker, which is the sinoatrial (SA) node in the right atrium. Despite the presence of any ectopic pacemakers, the SA node continues to fire, meaning that the cardiac muscle cells receive electric currents from more than one direction. The synchronized contraction of the heart, which begins in the atria and flows through the ventricles, is lost. If there is enough ectopic pacemaker activity, a condition called ventricular fibrillation may develop, in which heart muscle cells in different areas contract independently of one another. In ventricular fibrillation, all coordinated pumping activity of the ventricles is lost.
The defibrillator sends a large electric current of short duration through the heart, with the objective of repolarizing all of the cells at the same time. If defibrillation is successful, the SA node and the heart’s normal conduction system will resume control over depolarization of the heart after the cells have been “reset” by defibrillation.
CARDIAC conduction system
Even though cardiac muscle is autorhythmic it does need some sort of control to keep it beating at a steady rate. Areas of cardiac muscle cells have developed that initiate an impulse that moves through the myocardium, stimulating first the muscle cells in the walls of the atria to contract, then moving through the walls of the ventricles to cause them to contract. After each contraction the muscle cells relax. One cycle of atrial and ventricular contraction and relaxation is a cardiac cycle. The cardiac cycle produces one heartbeat.
The impulse for each heartbeat comes from the sinoatrial node (SA node) located in the wall of the right atrium. It is the pacemaker of the heart. The SA node is an area of cardiac muscle cells that automatically generate the impulses that trigger each heartbeat.
At rest, a cardiac muscle cell is polarized. Sodium and calcium ions are located on the outer membrane of the cell and potassium ions are located inside the cell. In order for the cell to contract it must depolarize. During cardiac muscle cell depolarization (Figure 14-16) two sets of events occur.
![]()
FIGURE 14-16 Depolarization and repolarization of a cardiac muscle cell. (From Wanamaker BP: Applied pharmacology for veterinary techinicians, ed 4, St Louis, 2008, Saunders.)
First, sodium (Na+) and calcium (Ca2+) ions move through channels in the cell membrane from the exterior to the interior of the cell. This reverses the polarity of the cell membrane. Second, potassium (K+) ions move through channels in the cell membrane from the interior of the cell to the exterior. This restores the original polarity, but now the Na+, Ca2+, and K+ ions are on the wrong sides of the cell membrane. During repolarization the ions are pumped back to their original locations and the cell is ready to depolarize again. In this way, the heart automatically keeps going through the cardiac cycle of cellular depolarization (contraction) and repolarization (relaxation).
The impulse generated by the SA node travels from the base of the heart to the apex and back to the base of the heart. The impulse passes through the muscle fibers of the wall of the atria. When the impulse passes through these cardiac muscles, the muscles contract. Remember, unlike other muscle fibers, cardiac muscle can transmit an impulse from one muscle cell to another, so impulses and the resulting muscle contractions spread across the atria in a wavelike fashion.
The structures that make up the primary cardiac conduction system are the SA node, the atrioventricular node (AV node), the bundle of His, and the Purkinje fiber system (Figure 14-17).
As stated above, after the impulse is initiated in the SA node in the right atrium, it spreads in a wave across both atria, causing them to contract and push blood through the AV valves into the ventricles, which are still relaxed. The impulse generated by the SA node travels through the walls of the atria to the atrioventricular node located in the atrioventricular septum that separates the left and right sides of the heart. The only route of conduction the impulse can take from the atria to the ventricles is through the AV node. When the impulse from the SA node reaches the AV node it delays
![]()
FIGURE 14-17 Cardiac conduction system. A, The SA node has initiated an impulse; B, the impulse has spread through the walls of both atria and reached the AV node (atrial depolarization); C, the impulse has traveled down the interventricular septum through the bundle of His, and the Purkinje fiber system has initiated ventricular contraction beginning at the apex, D, the impulse has spread through the walls of both ventricles (ventricular depolarization). (From Cunningham JG: Textbook of veterinary physiology, ed 4, St Louis, 2007, Saunders.)
for a fraction of a second. The delay permits the atria to complete their contraction before ventricular contraction begins. If atrial and ventricular contraction took place at the same time, the pressure in the contracting ventricles would be so high that the weaker, thin-muscled atria could not push blood into the ventricles.
After the delay at the AV node, the impulse resumes its speedy journey, this time through the bundle of His and the Purkinje fiber system. The fibers of the bundle of His travel down the interventricular septum to the apex of the heart. Here the Purkinje fiber system picks up the impulses, makes a U-turn, and carries them from the bundle of His up into the right and left ventricular myocardium. Because the impulse is delivered to the apex more quickly than it can spread from cell to cell in the ventricular myocardium, the ventricles actually contract starting from the apex and moving toward the base of the heart. This apex-to-base direction of ventricular contraction facilitates ejection of blood into the aorta and pulmonary arteries, which are located at the base of the heart (Box 14-1).
Just as the atria begin their contractions while the ventricles are relaxed, the atria also enter their relaxation phase while the ventricles are contracting. When the ventricles are contracting but the atria are relaxed, the pressure in the ventricles is much higher than the pressure in the atria, so
BOX 14-1 Try This At Home
As mentioned in the text the two atria contract while the two ventricles relax and vice versa. To get a better sense of how this works make a fist with both hands. Put your right fist representing the atria on top of your left fist representing the two ventricles. First squeeze your right fist in a rolling movement starting with your thumb and forefinger and going down. This represents the systolic wave of the atria, which begins at the cardiac base and moves to the atrioventricular septum. Now as you relax your right fist (atrial diastole) squeeze your left fist (ventricular systole) in a rolling movement starting with your little finger and moving up. The systolic wave through the ventricle starts at the cardiac apex with the Purkinje fiber system (i.e., your pinkie finger), and moves toward the base (i.e., your thumb). Relax your left fist and squeeze your right fist again. Each time you go through one squeeze and relaxation of each fist you have completed a cardiac cycle. Once you have mastered the squeezing and relaxing sequence, try and set up a rhythm of contractions of one “cardiac cycle” per second. This represents a heart rate of 60 beats per minute (bpm).
the AV valves are forced shut. With the AV valves closed, the relaxed and expanding atria can fill with blood again. At about the time the atria are becoming completely full, contraction comes to an end in the ventricles, and they begin to relax. This results in the pressure in the ventricles dropping lower than the pressure in the arteries they supply, so the aortic and pulmonary valves are forced shut. Pressure in the ventricles also falls below the pressure in the full atria, so the AV valves are pushed open.
After the AV valves open, the ventricles again fill with blood from the atria. The negative pressure caused by ventricular relaxation pulling blood in from the atria generates most ventricular filling. Just as the pressure in the atria and the ventricles begins to reach equilibrium, the SA node “fires,” causing the atria to contract and forcibly push the rest of their blood into the ventricles, and the cardiac cycle begins again.
There are two clinical terms associated with contraction and relaxation of cardiac muscles. Systole is the myocardium contracting, causing a chamber to empty itself of blood. This is the working phase of the cardiac cycle when the cardiac muscle cells are depolarized. Diastole is the myocardium relaxing and repolarizing after a contraction, allowing the chambers to fill with blood again. This is the resting phase of the cardiac cycle.
Each chamber goes through systole and diastole, but not all at the same time. In one cardiac cycle first the atria contract (atrial systole) while the ventricles are relaxed (ventricular diastole). This allows blood to flow from the atria into the ventricles. Next the ventricles contract (ventricular systole) while the atria are relaxed (atrial diastole). This allows the ventricles to eject blood from the heart and for the atria to fill with blood again.
NORMAL HEART SOUNDS
If you've had a chance to listen to a heart with a stethoscope you know it makes sounds as it beats. The heart valves snapping shut produce these sounds. One cardiac cycle produces two distinct heart sounds (Box 14-2). The sounds are often described as “lub” and “dub.” The first sound, “lub,” is produced when the tricuspid and mitral valves snap shut after atrial systole. The pressure in the ventricles is greater than the pressure in the atria at this point so the AV valves are forced closed. Remember these are one-way valves so blood cannot backflow into the atria.
The second heart sound, “dub,” is produced after ventricular systole when the pulmonary and aortic valves (semilunar valves) snap shut. The pressure in the pulmonary artery and aorta exceeds that in the ventricles at this point so the valves are forced closed. These are also one-way valves so blood cannot backflow into the ventricles. Most heart sounds are best heard on the left side of the standing animal by placing the stethoscope on the chest wall at about the point of the elbow. From this point, by moving slightly forward and backward, the pulmonary, aortic, and mitral valve sounds are heard. The tricuspid valve sound is best heard on the left side of the standing animal at approximately the same level (Figure 14-18).
![]()
![]()
FIGURE 14-18 Approximate locations for auscultation of the cardiac valves on the thoracic wall. T, Tricuspid valve; P, pulmonary valve; A, aortic valve; M, mitral valve. (From Nelson RC: Small animal internal medicine, ed 4, St Louis, 2009, Mosby.)
ABNORMAL HEART SOUNDS
If the two AV valves or the two semilunar valves are not closing simultaneously you may hear extra heart sounds.
Valvular insufficiency is a heart condition where one or more of the cardiac valves don't close all the way. When this happens a murmur is produced. The murmur sound is produced by turbulence in the blood flow and sounds iike a swishing or whooshing sound rather than a lub or dub. hi the case of valvular insufficiency the murmur is lcyoaoudsed b b backflowing abnormally into a chamber. For example, a murmur caused by mitral valve insufficiency results ⅛n the mitral valve doesn't close all tyhheen wa w the left ventricle begins systole. Instead of all the blood being ejected through the aortic valve into the aorta, some of the blood backflows into the left ahtirsium. T results in less blood entering the systemic circulation.
Valvular stenosis is a heart condition where any one or fmore o the cardiac valves don't open all the way. Again a murmur is produced by turbulent blood flow. In the case aolfvuvlar stenosis the murmur is caused by blood ofluogwhing thr a partially open valve and producing the same whooshing or swishing sound. For example, a murmur caused by mitral valve stenosis remits ⅛n the amlviteral v doesn't open entirely during left atrial systole. Instead of all the blood from the left atrium being ejected ifnt to the le ventricle, some of the blood remains in the lreiuftma.t This results in less blood in the left ventricle jteeoctbede into the systemic circulation. It also results in less blood being able to enter the left atrium from the pul- cmuolantaioryn cir because the left atrium doesn't empty
completely.
TEST YOURSELF 14-4
1. What isthepacemakerof the heart and where is it located?
2. List thcfeurconductorsthat mahe Uf the rapidconduc- tioe system fat ae impulse created by the heart's pacemaker.
3. Thewpuldng p ha caoh acarCie c cycle it. Iti nvolves
_______ that generates ae impulse that results ie muscle coetractioe.
4. What ij5gae pest ng er reemeart khambers
durieg left atrial diastole?
5. Wliee the m HralraIue isfaruedulouddit produces part of which heart sored, the first or the secoed?
CARDIAC OUTPUT
The cardiac output (CO) is the volume of blood that is ejected ou of the left ventricle over a unit of time, usually 1 nminute. I a healthy animal the cardiac output has to be soufficient t supply oxygen and nutrients throughout the animal's body. Two factors determine the cardiac output (1) stroke ^me and (2) heart rate.
CLINICAL APPLICATION
Congestive Heart Failure
In older dogs, congestive heart failure (CHF) is a fairly common CHF occurs when the pumping ability of
the heart decreases, usually due to disease of the heart muscle or a valve malfunction that restricts the forward flow of blood through a valve or allows a backward flow. CHF may be predominantly right-sided or left-sided. When the right fide of the heart begins to fail, blood returning from the scyusltaetmioinc cir is no longer able to move through the
right heart as quickly. This causes increased blood pressure isntemthiecsy circulation, which results in fluid accumulation in the ⅛m of ascites (fluid in the abdomen) and edema (fluid in the tissues).
When the left side of the heart fails, venous return from the I ungs is decreased, resulting in pulmonary edema, which interferes with respiratory function. The decrease in cardiac coiuatpedut asso with heart failure may also reduce perfu-
fsoirotnanot imp organs, such as the kidneys, to dangerously lvoewls.le
Medications used to treat CHF include cardiac glycosides teoasiencr the strength of cardiac contractions, diuretics to promote elimination of extra fluid to relieve edema, and vasodilators to enhance blood flow to the organs and decrease vascular resistance to outflow from the heart. CHF in pets ceannot b cured, but it can often be medically managed to improve the animal's quality of life.
The stroke volume (SV) is the ^me of blood ejected from the left ventricle during one contraction or systole. Another name for the stroke volume is systolic discharge. The heart rate (HR) is the number of times the ventricle rcontracts o beats in 1 minute. For an individual animal the HR is based in part on the rate at which the SA node spontaneously depolarizes:
cardiac output (CO) = stroke volume (SV) ? heart rate (HR)
For exemple if we know that a dog's heart ejects 100 milliliters (ml) of blood into the aorta with each systolic contraction, and its heart rate is 100 beats per minute, the animal's cardiac output can be calculated using our formula:
CO = 100 ml/min (SV) ? 100 beats/min (HR) = 10,000 ml/min
Cardiac output for this dog would be 10,000 ml of blood per minute. This equation can help explain why cows and horses istuhrvive w much slower heart rates than cats and dogs. A rhoworse o c needs more cardiac output than a dog or cat because it has much greater tissue mass, yet it normally has a slower heart rate. How can that be? Well, if
CO = SV ? HR
aonesd CO g up but HR goes down, we can write the equation as:
CO = ?SV ?T HR
In order to balance the equation the stoke volume has to increase. Horses and cows have much larger hearts than dogs and cats; this allows them to have such a large stroke volume that they can generate sufficient cardiac output, even with a slower heart rate than smaller animals.
The stroke volume represents the strength of the heartbeat. It is determined by two factors: preload and afterload. Preload is the volume of blood the ventricle receives from the atrium. Eighty percent of ventricular filling occurs passively by gravity; the remaining 20% happens during atrial systole when the atrium contracts. When the atrium doesn't contract sufficiently the preload will decrease and the ventricle receives less blood.
Afterload is the physical resistance presented by the artery the ventricle is ejecting blood into. If there is resistance in the artery (e.g., partial blockage) the ventricle will not be able to contract all the way and the amount of blood ejected into the artery will be decreased.
Another factor that can affect the stroke volume is the length of the cardiac muscle cells. The longer the cells the more force they produce when they contract. The myocardial cells can be stretched to a certain degree by introducing an increased amount of blood into the ventricle during ventricular diastole. In this way myocardial cells are somewhat elastic. So if the ventricular wall is stretched the ventricle chamber increases in size and holds more blood. In order to get rid of the increased amount of blood the ventricular wall has to contract with increased force. The result is an increased stroke volume.
The normal heart rate for each species of animal is set internally by the rate of spontaneous SA node depolarization (Box 14-3). External control of the heart rate comes through the autonomic nervous system (discussed in Chapter 9).
TEST YOURSELF 14-5
1. Stroke volume (SV) is a measurement of what?
2. If the cardiac output and stroke volume both decrease what has to happen to the heart rate to achieve equilibrium?
3. What is the difference between the preload and the afterload in reference to the stroke volume?
4. How could mitral valve stenosis affect the stroke volume?
| BOX 14-3 | Normal Heart Rate (and Pulse Rate) for |
| Common Animal Species | (*bpm) |
| Cat | 120-140 | Goat | 70-80 |
| Dog | 70-120 | Pig | 70-120 |
| Cow | 36-60 | Chicken | 250-300 |
| Dairy cow | 48-84 | Guinea Pig | 200-300 |
| Horse | 28-40 | Rabbit | 180-350 |
| Sheep | 70-80 | Rat | 250-400 |
| *Beats per minute. |
BLOOD VESSELS
Blood vessels in the pulmonary and systemic circulations make continuous loops to and from the heart. In the pulmonary circulation this means the blood vessels are branches from the pulmonary artery and vein. In the systemic circulation the blood vessels are branches from the aorta and vena cava. All blood vessels are arteries, veins, or capillaries. They are hollow tubes with similar but not identical anatomy and function.
The walls of arteries and veins have three layers (Figure 14-19). The inner layer that lines the lumen of the vessel is the endothelium. It is composed of thin, smooth simple squamous epithelium and is continuous with the endocardium that lines the chambers of the heart. The endothelium provides a smooth surface for the vessel lumen so blood flows easily through the vessel with little or no friction.
The middle layer of a blood vessel wall is made up of smooth muscle, elastic fibers, or both. The smooth muscles contract and relax to change the diameter of the vessel. They are controlled by the autonomic nervous system. The elastic fibers provide stretchability to the vessel wall. They allow the blood vessel to stretch and recoil without outside control.
The outer layer of a blood vessel is composed of fibrous connective tissue and collagen fibers. The connective tissue is strong and flexible which prevents vessel walls from tearing. The collagen fibers extend outward from the connective tissue and anchor the vessels so they can't move around too much. They also help keep the lumen of the vessel pulled open.
ARTERIES
Arteries carry blood away from the heart. In the pulmonary circulation they carry deoxygenated blood to the lungs for oxygenation. In systemic circulation they carry oxygenated blood throughout the animal's body. There are two types of artery: elastic arteries and muscular arteries. Elastic arteries have the greatest ability to stretch when blood passes through them because they have a large number of elastic fibers in the middle layers of their walls. These arteries are found closest to the heart because they have to be able to stretch and recoil without damage each time a surge of blood is ejected from a ventricle during ventricular systole.
The aorta is the largest elastic artery in the body. It has the largest layer of elastic fibers in its wall because it must be able to withstand the entire surge of blood ejected from the left ventricle. Other elastic arteries branch off the aorta. They have smaller diameters than the aorta but still have more elastic fibers than smooth muscle fibers in their wall.
Muscular arteries have more smooth muscle fibers than elastic fibers in their walls. They are found farther away from the heart than elastic arteries and usually direct blood to specific organs and tissues. Muscular arteries branch off the smallest elastic arteries and therefore have a smaller diameter. They are located far enough away from the heart that the blood surge is not severe enough to cause damage. Muscular arteries branch into arterioles.
![]()
FIGURE 14-19 Anatomy of arteries and veins. (From Colvile J, Oien S: Clinical veterinary language, St Louis, 2014, Mosby.)
Arterioles are the smallest branches of the arterial tree. They are in effect small muscular arteries and have the narrowest diameter. Blood flow to areas of the animal's body is controlled through contraction of the smooth muscles in their walls under autonomic nervous system control. Their smaller diameter produces blood flow resistance, which in turn helps maintain blood pressure.
Muscular arteries and some elastic arteries are frequently named for the organ or tissue they are carrying blood to (e.g., the renal artery carries blood to a kidney). An animal's body is pretty much bilaterally symmetric so arteries usually come in pairs (e.g., the right and left ovarian arteries supply blood to the right and left ovar y respectively) (Figure 14-20).
CAPILLARIES
Arterioles branch into many microscopic blood vessels called capillaries. Capillaries do not occur singly but in groups called capillary beds or capillary networks. One arteriole will give rise to an entire capillary bed. The wall of a capillary is one endothelial cell thick. It has no middle or outer layer. For this reason the exchange of gases and nutrients takes place at this level. No cell in an animal's body is far from a capillary bed so the waste products of cellular metabolism are easily exchanged for the oxygen and nutrients necessary for cellular metabolism.
VEINS
In order to get the blood back to the heart the capillaries join together to form tiny veins called venules. In the pulmonary circulation the venules carry oxygenated blood; in the systemic circulation they carry deoxygenated blood and waste materials. Venules have thin enough walls that some fluid exchange between interstitial fluid and plasma can take place. Their walls consist of endothelium, a thin muscle layer, and a few fibrous connective tissue cells. White blood cells leave the circulation at the venule level to enter tissues at a site of inflammation.
Venules join together to form veins. Veins and arteries in a specific area run close to each other so veins are named for their corresponding arteries. For example, the femoral veins that drain blood from the hind legs accompany the femoral arteries that supply blood to the hind legs.
As veins approach the heart they become larger in diameter as more veins draining other areas of the body join together. The largest vein in the animal's body is the vena cava, and all other systemic veins eventually drain into it. Figure 14-21 shows the major veins in a cat's body.
Many veins are working against gravity to get the blood back to the heart and they don't have the force of ventricular contraction to propel blood flow. For this reason small and medium veins have one-way valves in their lumens. The valves allow blood to flow only in the direction of the heart. When blood tries to flow backward, the valves close. These valves are similar to the semilunar valves in the heart, but they each have only two cusps. Muscular movements in the body compress small veins and the one-way valves allow blood to move only toward the heart. This is the only mechanism that propels blood back to the heart.
![]()
FIGURE 14-20 Major arteries of the cat. (Modified from McBride DF: Learning Veterinary terminology, ed 2, St Louis, 2002, Mosby.)
BLOOD CIRCULATION IN THE FETUS
Now that you are familiar with how blood moves through an animal, we'll look at some differences in blood flow in a developing fetus. The major difference between a fetus and a newborn is that the newborn receives oxygen through its own lungs, and a fetus receives oxygen from the blood of its mother. Because fetal lungs are not used for oxygen/carbon dioxide exchange, they need only enough blood to keep the growing lung tissues alive. Consequently, in the fetus there are bypasses that allow most of the blood in the fetal circulation to go around the lungs instead of through them.
The fetus receives oxygen through the placenta, an organ containing a network of tiny blood vessels that allows oxygen exchange between fetal and maternal circulation (Figure 14-22). The oxygenated blood from the mother flows from
![]()
FIGURE 14-21 Major veins of the cat. (Modified from McBride DF: Learning Veterinary terminology, ed 2, St Louis, 2002, Mosby.)
the placenta into the fetus through the umbilical vein. The vessel that carries oxygenated blood to the fetus is called a vein because it flows toward the heart of the fetus.
The oaye^^^ed blood in the umbilical vein flows through the fetal liver and into the caudal vena cava, where it mixes ewoixthygdenated blood from the fetal systemic circulation. Just as in the newborn animal, blood from the vena cava fills the right atrium. However, in the fetus two structures allow fmost o the fetal blood to bypass the lung tissue, because the blood in the right atrium has already been oxygenated from the maternal blood, and the lungs of the fetus do not perform oxygen exchange. The first bypass is the foramen ovale between the right and left atria (foramen means “opening,” and ovale means “oval”). Much of the blood from the right atrium flows directly into the left atrium, but some does flow through the tricuspid valve into the right ventricle and then
![]()
FIGURE 14-22 Circulation in the fetus and the newborn. A, Fetal circulation, B, newborn circulation. LA, Left atrium; LV, left ventricle; RA, right atrium; RV, right ventricle.
into the pulmonary artery. Blood from the pulmonary artery may flow into the lungs or through another bypass, the ductus arteriosus, directly into the aorta. Remember that this blood was oxygenated when it passed through the plocenta. Blood travels through the fetal aorta to the fetal systemic circulation, where it supplies oxygen and collects waste products from the tissues. The deoxygenated blood is then sent back to the placenta for oxygenation through the tuemriebsi.lical ar
With the first breath after birth, the lungs inflate, and the newborn begins to oxygenate its own blood. In a normal newborn, the foramen ovale and ductus arteriosus close at itmhaet t so that blood can no longer bypass the lungs.
TEST YOURSELF 14-6
1. From theαortic valve i n the IeO ventric Iesto the right atrium, list, ic order, the typos of blood eossol a drop of blood will pass through.
2. TOocvronarv a rθeryis VhrfitsO oaa not off^e a orta. Is it a muscular or olastic artory? Why?
a. Ic g ^egnaot ewe, whio h o rethe onl y two veins that aro carrying oxygenated blood?
4. Vbyat tws vre Oou adin the fetus that
allow most of its blood to bypass tho pulmocary circulatioc?
![]()
CLINICAL APPLICATION
Patent Ductus Arteriosus
During gestation, the fetal circulation does not carry much blood through the lungs; instead, there is an opening between the left pulmonary artery and the aorta that allows much of the b 1ood that leaves the right ventricle to bypass the lungs on the way to the aorta and systemic circulation. Because the fetus ⅛is not breathe air—oxygen is provided to the fetus from the blood of the dam—there is only enough pulmonary circulation to nourish the tissues of the growing lungs. Following the rupture of the umbilical cord at birth, the fetus emnuesrtateg its own oxygenated blood, so circulation through the lungs must be increased to include all of the blood that leaves the right ventricle.
Normally, the shortcut opening between the pulmonary artery and aorta, known as the ductus arteriosus, doses soon after feh. Occasionally, the opening fails to close in the newborn, a condition called patent ductus arteriosus (PDA). Yomg animals with PDA suffer from inadequate oxygenation of their blood; in the long term, the condition iosminpcatible with life. PDA may be treated with drug trohoemraoptey t p closure, or surgical closure of the ductus arteriosus may be an option.
PULSE
The pulse is the rate of alternating stretching and recoiling of the elastic fibers in an artery as blood passes through it with each heartbeat. The artery has to stretch and recoil because the left ventricle doesn't eject blood in a continuous flow. Every time the left ventricle contracts (systole) it ejects a bolus of blood into the aorta. When the left ventricle relaxes (diastole) blood flow into the aorta stops. This sets up a pulse wave of stretching and recoiling that travels through all the arteries and arterioles and dissipates in the capillaries. In most animals the pulse is felt on superficial arteries lying against firm surfaces such as bones.
Clinically the pulse is used to evaluate the regularity of the pulsations and the strength of the pulsations. The pulse and the heartbeat in an animal are not the same thing. The pulse is felt on a superficial artery; the heartbeat is counted using a stethoscope to listen to the animal's chest (auscultation) to hear the heart sounds. In some animals the heartbeat can be felt through the chest wall. This is still not a true pulse because it is not felt over an artery.
PULSE POINTS
The pulse is best felt on different arteries in different species of animal (Box 14-4). It is most commonly evaluated over a medium-sized artery. In a healthy animal the pulse rate and the heart rate should be equal. When feeling for a pulse, use the tips of your index and middle fingers, not your thumb because the thumb has its own pulse (Figure 14-23). The pulse wave that passes through your thumb may be confused
| BOX 14-4 | Common Pulse Points in Common Animal Species |
| Cat | Femoral artery—medial surface of the thigh near the belly |
| Dog | Femoral artery—medial surface of the thigh near the belly |
| Cow | Coccygeal artery—ventral midline of the tail near the base Facial artery—passes over the mandible near the angle of the jaw |
| Horse | Mandibular artery—passes over the mandible near the angle of the jaw Posterior digital artery—on the pastern between the coronary band and the fetlock |
| Sheep | Femoral artery—medial surface of the thigh near the belly |
| Goat | Femoral artery—medial surface of the thigh near the belly |
| Piglet | Femoral artery—medial surface of the thigh near the belly |
| Pig | Coccygeal artery—ventral midline of the tail near the base |
with the animal's pulse. As a general rule, larger animals have slower pulse rates and smaller animals have faster pulse rates. The pulse cannot be taken over a vein because the pulse wave dissipates in the capillaries and none of it is passed into the veins.
BLOOD PRESSURE
As the name implies blood pressure is a measure of the amount of pressure flowing blood exerts on arterial walls. It is dependent on the interaction between the heart rate, stroke volume, the diameter and elasticity of the artery, and the total blood volume. Any condition or medication that affects any one or more of these factors will also affect the blood pressure.
![]()
FIGURE 14-23 Finding the pulse on A, a dog or cat, B, a cow, C, a horse. (A, From Thomas J, Lerche, P: Anesthesia and analgesia for veterinary technicians, ed 4, St Louis, 2011, Mosby; B and C, From Bassert J, McCurnin D: McCurnin's clinical textbook for veterinary technicians, ed 8, St Louis, 2014, Saunders.)
Blood pressure varies during the cardiac cycle. If you’ve ever had your blood pressure measured you know that two numbers are recorded. One number, the highest number, is the systolic blood pressure. It is produced by ejection of blood from the left ventricle into the systemic circulation by way of the aorta. The second number, the lowest number, is the diastolic blood pressure. It measures the pressure remaining in the artery during left ventricular diastole when the ventricle is relaxing and refilling with blood. A third value, the mean arterial pressure (MAP), is sometimes measured. This is the average pressure during one cardiac cycle. The MAP can be used when monitoring an anesthetized animal as an indication of tissue perfusion.
Blood pressure is most commonly measured using one of two methods. The oscillometric method is the method often used in your doctor’s office. A cuff is placed over the area of an artery and inflated until blood flow either stops or nearly stops. The air is gradually released from the cuff and an instrument attached to the cuff measures the magnitude and frequency of pulsations. The instrument automatically determines the systolic and diastolic blood pressures. Many instruments will also determine heart rate.
The second method used to determine blood pressure is Doppler ultrasound. The ultrasound instrument measures arterial blood flow as air is released from an inflated cuff attached to a sphygmomanometer. The cuff is proximal to the position of the Doppler transducer that is placed over an artery. This method accurately measures systolic blood pressure only.
CARDIOVASCULAR MONITORING
Given that the heart is not visible by direct observation, a number of direct and indirect tests have been developed to monitor it and the entire cardiovascular system. These tests include:
• Auscultation of the thorax to determine heart rate and rhythm, and to detect heart murmurs
• Peripheral artery palpation to evaluate the rate, regularity, and strength of the pulse
• Measurement of arterial blood pressure to evaluate cardiac output
• Thoracic radiography to evaluate the size and position of the heart
• Electrocardiography to evaluate the electrical activity of the heart
• Echocardiography to evaluate the size, shape, and movement of heart structures
ELECTROCARDIOGRAPHY
Electrocardiography produces an electrocardiogram (ECG or EKG) based on the electrical activity of the heart. The electrical impulse that originates in the SA node and spreads through the cardiac conduction system can be detected on the surface of the animal’s body. Leads are placed at specific locations on the animal’s body and attached to an ECG machine. When the ECG machine is turned on, the electric impulses generated in the heart muscle are picked up at the
![]()
FIGURE 14-24 The electrocardiogram (ECG) of normal heartbeats. The enlarged area represents one cardiac cycle or heartbeat. (Adapted from Koeppen BM, Stanton BA: Berne & Levy physiology, ed 6, St Louis, 2010, Mosby.)
various locations on the body surface where the leads were placed, and are recorded. A series of heartbeats are followed for a specified period of time, recording them at a standardized rate on a strip of graph paper moving through the instrument.
Figure 14-24 shows the ECG of one cardiac cycle. An explanation of the various components in Figure 14-24 follows:
• P wave
• Is the time it takes the wave of depolarization (contractions) to travel from the SA node through the atria
• It corresponds to the mechanical activity of atrial contractions in a normal animal
• QRS complex
• Is the time of ventricular depolarization (contraction)
• It corresponds to the mechanical activity of ventricular contraction
• It is composed of three different waves
• Q wave corresponds to depolarization of the interventricular septum.
• R wave corresponds to depolarization of the main mass of the ventricles so it is the largest wave
• S wave corresponds to the final part depolarization of the ventricles near the base of the heart
• T wave
• Is the time of ventricular relaxation (repolarization)
• It corresponds to the time taken by the ventricles to get ready for the next contraction by refilling with blood from the atria
ECHOCARDIOGRAPHY
Another way to evaluate the heart is through echocardiography (ECHO or cardiac ultrasound). This procedure uses ultrasound to bounce sound waves off parts of the heart to watch the heart beating. ECHO can be used to evaluate the size, shape, and movement of the heart and its parts.
Two-dimensional echocardiography produces an overall 2-D cross-sectional image of the heart (Figure 14-25). (The image is much like the image produced of a fetus when ultrasound is used on a pregnant woman.) Two-dimensional echocardiography is a trans-thoracic procedure where the
![]()
FIGURE 14-25 Two-dimensional echocardiography (ECHO) of a heart with an interatrial septal defect (A). All four chambers of the heart are visible. (Modified from Ettinger SJ, Feldman E: Textbook of veterinary internal medicine, ed 7, St Louis, 2010, Saunders.)
![]()
FIGURE 14-26 Doppler echocardiography showing blood flowing between the two atria (B) of the same heart as in Figure 14-25. (Modified from Ettinger SJ, Feldman E: Textbook of veterinary internal medicine, ed 7, St Louis, 2010, Saunders.)
transducer that emits ultrasound waves is moved over the thorax in the area of the heart. The sound waves bounce or “echo” off the heart and are sent to a computer that produces an animated image of the heart. Two-dimensional echocardiography is especially useful for evaluating the relative size of the heart chambers, the thickness of the myocardium, and the functioning of the valves. With this method you are able to see the heart working in real time.
Doppler echocardiography (Figure 14-26) is a more sophisticated ultrasound procedure that measures blood flow through the heart and adds color to the image. It is usually done in conjunction with two-dimensional echocardiography and is especially useful for evaluating valvular stenosis and insufficiency.
VENIPUNCTURE
Most blood vessels are buried deep within an animal's body, but some are superficial and can be seen or felt just under
![]()
the skin. These superficial blood vessels are used to collect blood samples, administer medications intravenously, and place venous or arterial catheters. It is important to become familiar with the names and locations of these superficial blood vessels (Figure 14-27).
In dogs and cats, the cephalic vein of the thoracic limb is commonly used for venipuncture. It runs between the elbow and the carpus on the cranio-medial aspect of the forearm. The cephalic vein carries blood from the distal extremity to the jugular vein.
In the pelvic limb of dogs and cats, the femoral vein or the saphenous vein may be used for venipuncture. The femoral vein, which is more commonly used in cats than Uogs, runs along the medial aspect of the hind limb between the groin and the tarsal joint (hock) and carries blood to the iliac vein, which joins the vena cava. The saphenous vheicinh, w is more commonly used in dogs, runs along the leactteral asp of the hind limb from the cranial aspect of the luoevsgte j ab the hock (tarsus) to the caudal aspect just below the knee (stifle). The saphenous vein carries blood to tehmeofral vein.
The jugular vein is commonly used for venipuncture in nearly all animal species, both large and small. Jugular veins tursacvuellairn m jugular grooves along the ventral aspect ceohf ea sid of the neck, from the mandible to the shoulder. The jugular veins are located near the carotid arteries, hi all suptecies, b especially in the horse, care must be taken to cacviodiedntaal injection into the carotid artery of sub
stances that are meant to be injected into the jugular vein. The carotid artery carries blood very quickly to the brain; substances that act as sedatives when normally injected into the l agni ar vein may actually cause seizures and/or death if accidentally injected into the carotid artery.
In Iantating dairy cattle, the superficial caudal epigastric vein, commonIy called the milk vein, is easily sαn as it travels eanlotnragl the v aspect of each side of the abdomen from the udder to about the level of the sternum. This vein is not to be used for routine venipuncture, because its thin-walled, shuapraecrtfiecrial c makes it prone to excessive bleeding and hematoma formation, which may lead to the development eosfsa. n absc
In ruminants and rodents, the eoccygeal vein may be eounrseipdufnctvure. The coccygeal vein carries blood from
tohe tail t the vena cava. It runs along the ventral midline of the tail.
TEST YOURSELF 14-7
1. What is the difference betwee n a heart ra te and a pulse?
2. What does systolic blooC psessure measure?
3. What iothedifference aetwaeh anECGand an ECHO?
4. Which sein cnn bessedinmaht eornmon animal speeies for venipuncture? Where is it located?