Metabolism and energetic definitions
We have all heard the expressions, "there are no free rides" or "no free lunches." This is certainly true when it comes to living our lives but is also a physiological
truth.
For example, acquiring the energy to maintain the ionic gradients we discussed in Chapter 2 depends on the hydrolysis of one of the phosphate bonds of adenosine triphosphate (ATP). The question becomes, "where did the energy needed to produce the ATP in the first place come from?" This is the crux of the problem; some molecules are Catabolized so that others can be created. Simply put, living systems are in a constant thermodynamic battle. As the second law of thermodynamics indicates, the natural tendency is toward equilibrium with dispersion of energy and increasing disorder, or in thermodynamic terms, increased entropy. Living systems, cells, tissues, organs, systems, and organisms are characterized by just the opposite. The degree of complexity and organization in living systems is antithetical to this law. Unlike most of the nonliving universe, open biological systems are able to exchange matter and energy with their surroundings. This allows living systems, while they are alive, to move away from the dispersion and energy equilibrium that the second law of thermodynamicsdictates. The first law of thermodynamics is familiar as the maxim that energy can neither be created nor destroyed. More formally, it is expressed in this way: the total energy of a system plus its surroundings remains constant. Thus, the exquisite organization and complexity that characterize living systems are at the expense of free energy from the environment; that is, energy that can be harnessed to do work. Thus, living systems are analogous to an oasis in the desert. The oasis, often short-lived in a geological sense, provides a place for relief for the weary traveler.
Living systems represent transient conditions during which time nonequilibrium conditions related to energy-matter circumstances exist.A fundamental postulate of theoretical biology is that life processes can be explained in terms of chemistry and physics—in other words, in terms of matter and energy. Building on this idea, it can be reasoned that life processes are represented by the myriad of chemical (enzymatic and nonenzymatic) and physical reactions that occur within cells and tissues—in other words, metabolism. The yin and yang of metabolism are anabolism and catabolism. Our discussion begins with catabolism.
The phrase "intermediary metabolism" is often used in nutrition and biochemistry texts. This refers to the many steps of reaction between the initiation of a biochemical process and its completion. For example, the complete oxidation of the critical nutrient glucose begins with uptake of the glucose into the cell and entry of the molecule into a sequence of reactions called glycolysis. This is an example of a biochemical pathway. The steps in the pathway detail the reactions that are required to convert this 6-carbon hexose sugar into two 3-carbon molecules of pyruvate. This process is also called anaerobic respiration. The various molecules that are temporarily produced in the 10 steps of glycolysis are called intermediates. Other biochemical pathways would generate their own specific family of intermediate molecules. Consequently, intermediary metabolism refers to the creation and existence of the hundreds of molecules that are fabricated as various molecules progress toward their final biochemical destination. Finally, although we typically describe important biochemical pathways singly, it needs to be emphasized that a host of biochemical reactions or pathways are occurring simultaneously. Intermediates from one pathway often also supply materials that can be used in other pathways. For example, one of the intermediate steps in glycolysis produces triose phosphate.
This molecule can supply the carbon atoms to make pyruvate or alternatively be shuttled out of the glycolysis pathway to produce glycerol that is needed in the anabolic pathway to make triglycerides. A key idea is that regulation and control of the rates of activity of these various, often competing biochemical pathways is critical. Resources must be used effectively and efficiently.Given the importance of energy in fueling the biochemical reactions in cells, it is fitting that we consider some definitions. What do we mean by energy? Are all forms of energy equally valuable from a physiological viewpoint? We often say that the diets we supply to our animals give them the energy they need for productive functions—for example, to allow draft horses to pull wagons, thoroughbreds to race, or cows to produce milk. Our animals do not consume energy directly, but rather it is the digestion of foodstuffs that liberates the nutrient molecules that can then be oxidized in a controlled, deliberate fashion to provide energy needs. In physics, energy is described as the capacity to do work. In biological systems, energy is sometimes expressed as heat units or calories. A calorie (cal) is the amount of energy needed to increase the temperature of 1 g of water 1 degree at a pressure of latm. Other related terms are the kilocalorie (1000 calories) or the megacalorie (Meal = 1,000,000 cal) often used in descriptions of the energy content of animals feeds. Other measures of energy are also frequently used. For example, IJ is the work done by a force of IN working over a distance of Im (m2 ? kg ? sec 2). Other measures of energy can be derived as well (leal = 4.187J or 0.004 British thermal units [BTU]).
It is also clear that not all of the bond or chemical energy that is available in food molecules can be captured for use by tissues or cells. If a known quantity of a food material or nutrient is completely combusted or oxidized in the presence of oxygen and the heat generated measured, this provides an estimate of the gross or potential energy of the substance.
This process is called calorimetry. This technique is valuable because it provides a measure of the potential energy available in a particular nutrient or other substance. However, as another old expression says, "it takes money to make money"; there are biochemical costs involved in acquiring the energy that is potentially available in various nutrients. Just as there are overhead costs in running a business or a university for that matter, there are physiological costs that must be paid to capture the energy available in nutrients. For example, consider feeding your horse a carrot and the steps needed to capture the energy in the starch. For processing there is mastication and swallowing, transport to the small intestine, secretion of gastric and intestinal enzymes, and transport of digested glucose molecules across the epithelial cells. All of these events can be thought of as overhead or maintenance costs associated with the physiological processing of the carrot.Thus, the amount of energy present in the diet does not equal energy that is ultimately available to the animal. Animal science nutritionists often conduct feeding trials to evaluate the practical value of different feedstuffs. These feedstuffs are often complex mixtures. Consider total mixed rations that are often fed to dairy cows with various combinations of forages and concentrates. In such studies, great care must be taken to account for measurement errors, for example, spilled feed and variation in the efficiency of digestion and ingestion. Particularly for complex rations, the gross energy content of the diet gives only a broad indication of how valuable the diet is to the animal. What happens if one of the components in the diet cannot be digested? What if there are interactions so that the breakdown of one dietary component affects the microorganism population of the GI tract so that normally effective nutrients are lost? Even under the best conditions, especially for animals fed fibrous feeds, a part of the feed is not digested and contributes to the energy content of the feces.
The difference between the gross energy content of the diet and the portion of the energy that is available to the animal is called digestible energy. Various rations can then be compared based on their digestibility energy, often expressed as a fraction or percentage of the gross energy. From measurements made over the course of several days and sometimes weeks, average daily energy consumption minus the energy content of the feces and urine is called the apparent digestible energy. It is called apparent because some generally small portion of fecal energy comes from sloughed intestinal epithelial cells, bacteria cells, and substances that have been excreted by way of the feces.For nutrients that are ultimately absorbed, the energy content still does not match energy that ultimately becomes available to the animal. However, the fraction that is represented by digested and absorbed nutrients is called metabolizable energy. This is the fraction of total or gross energy fed that is directly available to the tissues and cells to be processed. This energy can be used for maintenance, for heat generation (sometimes considered a waste product), or for more recognizable practical productive functions, that is, muscle growth, egg production, or milk production. Metabolizable energy is less than digestible energy, typically about 80%, because of other losses in addition to the fecal losses. This fraction would be lower still if the heat that is generated is considered a waste product. The additional losses include materials that are lost in urine as well as gaseous product from gut fermentation that are expelled. These products are especially plentiful in herbivores and particularly ruminants. The fraction of energy that is ultimately used for physiological activities is called net energy. Figure 3.1 illustrates the processing of dietary energy.
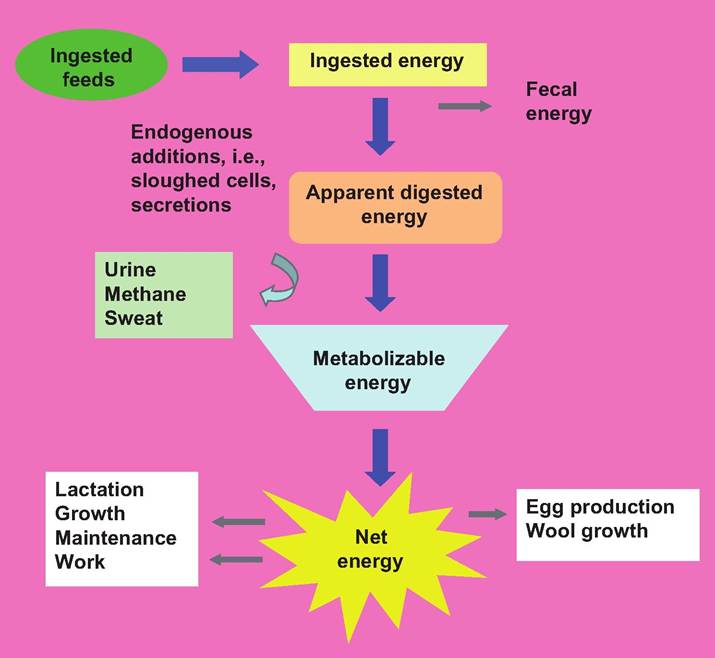
Fig. 3.1. Processing of feedstuff energy.
Compared with the total estimate of gross energy in consumed feeds, the amount of energy ultimately available to cells and tissues is substantially reduced.It is common to characterize energy that is needed for basal or resting life activities in animals as the energy of maintenance. These energy costs do not reflect energy that is needed for productive work in a production agriculture sense—for example, work done by a draft horse or racehorse or energy recovered from products such as meat, milk, or eggs. These are critical physiological functions, action of the Na-K ATPase pump, cardiac function, and so forth. As Figure 3.1 illustrates, there are additional energy costs associated with acquiring feed and assimilating nutrients themselves. Ideally, maintenance activities are stated as the energy costs of preserving an adult animal under resting or sedentary circumstances, in the absence of weight gain or loss, in thermoneutral conditions. In practical terms, this ideal status is rarely obtained, but it does give a framework to understand the meaning of maintenance energy costs. In humans, the term basal metabolic rate may be more familiar and is determined for subjects in a quiet, thermoneutral environment, about 12 hours after a meal.
Basal energy needs are determined indirectly by measuring oxygen consumption under these conditions. This is effective because the amounts of oxygen needed to completely oxidize fats, proteins, or carbohydrates are known. If CO2 production is simultaneously measured, the respiratory quotient can be calculated (moles of CO2 exhaled divided by the moles of O2 consumed). These data can then be used to estimate the nature of the nutrients (protein, carbohydrate, or fat) being oxidized for energy production. The alternative to indirect calorimetry is to measure heat production along with completed collection of gases and other wastes. This is clearly a difficult and expensive undertaking with large domestic animals. However, Table 3.1 shows an example of calorimetry data from a study by Tyrrell et al. (1988) designed to determine in part if the metabolic rate of dairy cows treated with bovine somatotropin (bST) was increased compared with controls. Briefly, nine cows received bST (51.5 IU/day) or a daily control injection in a single reversal experimental design, which utilized 14-day treatment periods. With increased milk production after bST (22% increase), cows already in a negative tissue nitrogen balance (-21 g/day) tended to become more negative during the bST treatment period (-34g/day). Energy and nitrogen balances were measured in open-circuit respiration chambers. As predicted from increased milk production, there was a corresponding greater heat energy loss and increased milk energy secretion after bST treatment. Tissue energy balance was -1.1 Mcal/day during the control treatment period. Increased use of energy reserves with bST treatment decreased tissue energy balance to -9.8 Mcal/day. These researchers concluded that much of the effect of bST to increase milk produc-
Table 3.1. Effect of bovine somatotropin (bST) in lactating dairy cows on milk production and energy metabolism parameters.
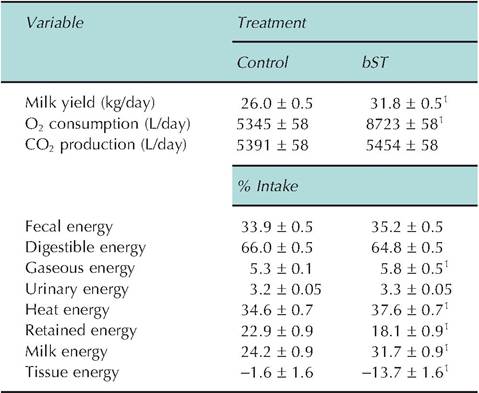
’Indicates a statistically significant difference between treatments
P < 0.5.
Data are adapted from Tyrrell et al. (1988).
tion in dairy cows is related to increased use of tissue reserves and altered partitioning of nutrients rather than dramatic effects on digestibility of nutrients or apparent changes in maintenance requirements of the animals. Simply, the increases observed would have been expected with the degree of increased milk production, regardless of the specific reason for increased production.