Metabolism of skeletal muscle
The major energy source for muscle metabolism is ATP. It is used for cross bridge formation, to actively pump calcium into the SR, and to pump Na+ out of and K+ into the muscle fiber.
The endogenous stores of ATP are able to last only about 4-6 seconds. Therefore, there must be mechanisms to replenish this limited store.While at rest, skeletal muscle makes sufficient ATP to meet its metabolic needs, and to store surplus energy in the form of creatine phosphate and glycogen. Resting muscle can use fatty acids that are broken down in the mitochondria and the ATP used to make creatine phosphate. The glucose that is delivered through the bloodstream can be converted to glycogen. When the demands for ATP become greater, there are three pathways for generating ATP: (1) aerobic respiration, (2) ADP interacting with creatine phos-
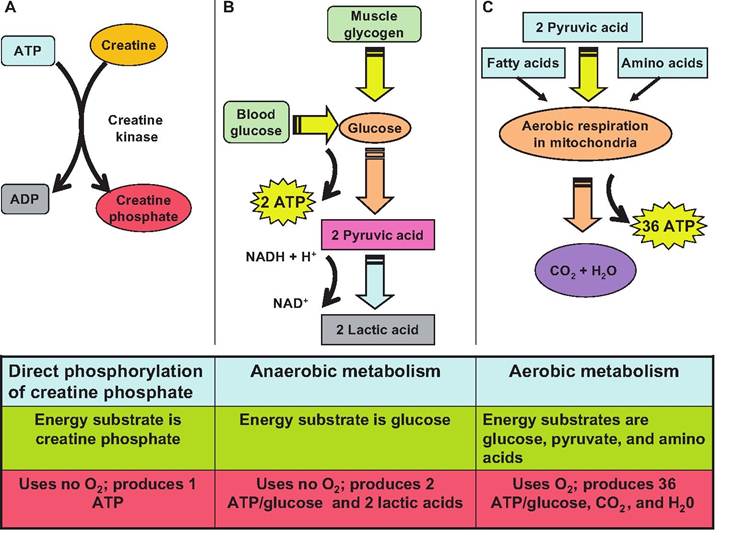
Fig. 7.18. ATP production in muscle. There are three mechanisms for generating ATP in muscle fibers. (A) During the direct phosphorylation of creatine phosphate, a phosphate group is moved from ATP to creatine producing creatine phosphate, an energy storage form in muscle. When ADP is plentiful, this reaction is reversed to produce ATP. (B) During glycolysis, glucose is anerobically broken down to two molecules of pyruvate, which are then converted to lactic acid in order to regenerate NAD+. (C) In the presence of O2, pyruvate is further metabolized to CO2 and H2O, plus ATP.
phate, and (3) from stored glycogen through the anaerobic process of glycolysis (Fig. 7.18).
Aerobic mechanism
When O2 is present, pyruvate enters the mitochondria where aerobic respiration occurs. This process produces 36 moles of ATP for every 1 mol of glucose.
During aerobic respiration, the following reaction occurs:Glucose + O2 → CO2 + water + ATP.
Since muscle is able to store glycogen, the breakdown of glycogen can yield glucose for aerobic metabolism. In addition, muscle can get energy from blood-borne glucose, pyruvic acid, free fatty acids, and some amino acids. During moderate exercise, aerobic metabolism is able to meet the energy demands of the muscle.
Creatine phosphate
While at rest, muscles are able to produce more ATP than is utilized. This surplus energy is used to synthesize creatine phosphate, an energy storage form found exclusively in muscle. The enzyme creatine kinase catalyzes the transfer of a high-energy phosphate group from ATP to creatine. When ADP is present in the muscle, creatine kinase catalyzes the transfer of a high-energy phosphate group from creatine phosphate to ADP forming ATP Creatine phosphate is three to six times more plentiful in muscle than ATP These two compounds together can provide enough energy for muscles to contract for approximately 10-15 seconds. When energy is plentiful, creatine phosphate is replenished.
Anaerobic mechanism
Although ATP and creatine phosphate can provide energy for short periods of time, other mechanisms are needed to produce ATP Glucose can be metabolized through glycolysis, the initial phase of glucose respiration. Glycolysis does not require oxygen, and therefore is termed an anerobic (without oxygen) process, anaerobic glycolyis. Therefore, this process is utilized when there is insufficient O2 available for aerobic metabolism to sustain the energy needs of the muscle.
During glycolysis, one molecule of glucose is metabolized to 2 molecules of pyruvic acid. During this process, 2 ATPs are produced per molecule of glucose. Therefore, anaerobic glycolysis can produce only 5% of the amount of ATP from glucose as aerobic respiration whereas aerobic respiration produces 95% of the ATP.
In the process of breaking down glucose, NAD+ is reduced to NADH + H+.
In order to continue glycolysis, the cell must be able to oxidize NADH back to NAD+. When oxygen is not present, the cell can do this only by converting pyruvate to lactic acid, during which NAD+ is regenerated. Lactic acid then diffuses out of the cells and goes to the liver, heart, or kidney. In the liver, it can be converted back to glycogen.Muscle fatigue and oxygen consumption
Muscle fatigue is the inability of muscle to contract after prolonged activity. This effect is due to conditions within the muscle fiber, since it occurs even in the presence of sustained neural input. Muscle fatigue differs from central fatigue, in which the muscle is still able to function but the mind is unwilling.
While the exact mechanism of muscle fatigue is unknown, several factors probably play a role. These include inadequate release of Ca2+ from the SR, depletion of creatine phosphate, insufficient O2, depletion of glycogen, buildup of lactic acid (decrease in pH) and ADP, or depletion of ACh from motor neurons. Furthermore, the lack of ATP in fatigued muscle may compromise the action of the Na+-K+ pump on the Sarcolemma, causing a loss of intracellular K+.
During prolonged exercise, muscles accumulate lactic acid. This lactic acid must be reconverted to pyruvic acid. In addition, the animal must replenish glycogen stores and replace creatine phosphate and ATP, and the liver must convert blood lactic acid to glycogen. These processes all require O2, leading to what was termed the "oxygen debt," or the amount of extra O2 that an animal must inspire to restore homeostasis. Since body temperature and enzymatic reaction rates are elevated for a period of time after exercise, a better term for the increased oxygen needed following exercise is recovery oxygen uptake. During the recovery period, the muscle fibers return to their pre-exertion conditions.
Heat production
Since metabolism is not 100% efficient, the by-product of metabolism is heat. While a resting muscle fiber is approximately 42% efficient, an active fiber is only about 30% efficient. The remainder of the energy appears as heat that is used to warm the tissues and fluids and help maintain normal body temperature. The excess heat produced when muscles are active is the reason body temperature climbs during exercise.
Types of muscle fibers
There are three functional ways to classify skeletal muscle fibers: (1) speed of contraction, (2) metabolic pathways forming ATP, and (3) myoglobin content:
1. Speed of contraction. Based on rate of contraction, fibers can be classified as fast or slow. Speed is based on the rate at which the myosin ATPase splits ATR
2. Metabolic pathways forming ATP Fibers that rely on oxygen-requiring pathways for generating ATP are called oxidative, whereas those that rely on anaerobic glycolysis are called glycolytic.
3. Myoglobin content. Some muscle fibers have a large amount of myoglobin, the red-colored protein that binds oxygen in muscle fibers. These fibers are termed red muscle fibers. In contrast, some fibers have low myoglobin content and are called white muscle fibers.
Based on these criteria, there are now three major categories of muscle fibers, including slow oxidative fibers, fast oxidative-glycolytic fibers, and fast glycolytic fibers—sometimes called slow, intermediate, and fast fibers, respectively. To further complicate matters, these fibers are also called Type I, Type II-A, or Type II-B fibers, respectively. While there are many more types of fibers, these are three such classifications that allow for the discussion of general characteristics (Table 7.1). For example, slow fibers allow for prolonged, sustained activities that are powered by aerobic metabolism and that fatigue slowly. Such fibers have a good blood supply, a high myoglobin content and many mitochondria. Conversely, fast glycolytic fibers are better suited for quick, powerful movements that occur over a short period of time and that fatigue quickly.
Since these latter fibers contract quickly, they rely on endogenous glycogen stores rather than glucose delivered via the bloodstream, and they have fewer mitochondria and little myoglobin.Effects of exercise on muscles
Exercise results in changes in the muscle. With endurance exercise, there is a prolongation of the time to fatigue. This resistance to fatigue appears associated with increases in aerobic power and anaerobic capacity. Such changes include increases in muscle buffering capacity and citrate synthase activity and reduced glycogen utilization. There is also an increase in the capillary-to-muscle fiber ratio, the number of mitochondria, and the amount of myoglobin.
With sustained but relatively weak muscle activity, the muscle does not display significant hypertrophy. However, high-intensity anaerobic activity is associ-
Box 7.1 Carnitine and exercise
Carnitine is needed for optimal mitochondrial oxidation. An animal gets carnitine from dietary sources such as meat or dairy products, endogenous biosynthesis, or from dietary supplements. While supplemental carnitine does not appear to enhance exercise performance in untrained humans, there is evidence that it may be beneficial in trained individuals. In recent studies conducted with Standardbreds, longterm (5 weeks) dietary supplementation with carnitine given in conjunction with exercise increased the percentage of type II-A fibers, increased the Capillary- to-fiber ratio, and appeared to spare muscle glycogen. ated with muscle hypertrophy. This is because this type of exercise is associated with strength more than with endurance. Such hypertrophy is due to increased muscle fiber size rather than hyperplasia (i.e., increase in cell number) (Box 7.1).