Contraction of skeletal muscle
As summarized in Figure 7.7, control of skeletal muscle contraction involves the voluntary stimulation of motor neurons innervating the muscle. The release of the neurotransmitter from these motor neurons initiates excitation-coupling-contraction in which an action potential is generated within the skeletal muscle fiber.
The action potential causes the release of calcium from the SR, which then causes muscle contraction.Neuromuscular junction
Skeletal muscle is controlled by the somatic nervous system. The cell bodies of the α-motor neurons, that is, somatic motor neurons, that innervate skeletal muscle reside in the central nervous system (CNS). The axons of these neurons leave the CNS and innervate skeletal muscle fibers at a specialized junction called the neuromuscular junction (NMJ) or myoneural junction (Fig. 7.8).
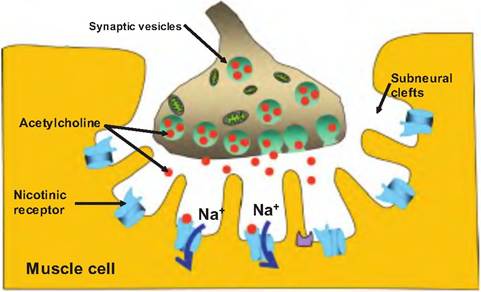
Fig. 7.8. Neuromuscular junction. The neuromuscular junction is a specialized synapse between an α-motor neuron and skeletal muscle fibers. The synaptic bouton is imbedded in the sarcolemma. At this site are subneural clefts that increase the surface area surrounding the synapse. Synaptic vesicles containing acetylcholine (ACh) are located in the nerve ending. Upon stimulation, the α-motor neuron releases ACh that can diffuse across the synaptic cleft and bind to nicotinic receptors on the skeletal muscle fiber. Upon binding to the receptor, ACh causes Na+ ions to enter the skeletal muscle fiber, causing a postsynaptic potential. The postsynaptic potential is always large enough to induce an action potential in the skeletal muscle fiber.
Each muscle fiber is innervated by a neuron, although a single neuron may innervate multiple muscle fibers. The neuron branches as it enters the perimysium, and each branch ends in a synaptic terminal, sometimes called a synaptic bouton.
The synapse is the region of contact between a neuron and its target cell, which in this case is a skeletal muscle fiber. The space between the neuron and the muscle fiber is the synaptic cleft. The Sarcolemma in the region of the NMJ is called the motor end plate.Since an electrical signal cannot traverse the synaptic cleft, the signal from the motor neuron is communicated via the release of a neurotransmitter. The neurotransmitter released from α-motor neurons is acetylcholine (ACh). ACh is contained in synaptic vesicles located in the synaptic bouton. When the action potential arrives at the synaptic bouton, it causes the release of ACh that then diffuses across the synapse and binds to a specialized cholinergic receptor located on the muscle fiber. This receptor is called a nicotinic receptor. This is a transmembrane protein that binds ACh, and can also be stimulated by the agonist nicotine.
When ACh binds to the nicotinic receptor, it causes the opening of a ligand-gated ion channel that allows sodium ions to enter the muscle fiber. This causes the production of a postsynaptic potential that results in the production of an action potential in the muscle fiber. The amount of neurotransmitter released per nerve impulse is greater than the amount needed to induce a postsynaptic action potential, and the number of receptors activated by ACh is more than required to reach threshold, thus leading to a "safety factor" assuring that stimulation of a motor neuron results in contraction of skeletal muscle.
Pharmacology of the neuromuscular junction
Since the NMJ is a chemical synapse, it is prone to pharmacological manipulation. Curare, produced by certain frogs, is a compound used by South American Indians to make poisonous arrows and darts. Curare blocks nicotinic receptors and thereby prevents ACh from inducing skeletal muscle contraction. Derivatives of curare are sometimes given prior to surgery in order to relax the skeletal muscles.
Clostridium botulinum is a bacteria often found in contaminated canned foods.
The toxin from this organism prevents the release of ACh from somatic motor neurons. Botulinum toxin prevents skeletal muscle contraction. A very tiny amount of this toxin can cause death by paralyzing the diaphragm and other respiratory muscles. Recently, this toxin has been increasingly used in human medicine (Botox) to reduce wrinkles, control strabismus (crossed eyes), blepharospasm (uncontrolled blinking), or cervical dystonia (also known as spasmodic torticollis), which is characterized by involuntary tonic contractions or intermittent spasms of the neck muscles.ACh is normally inactivated by the enzyme acetylcholinesterase (AChE). Agents known as AChE inhibitors can be used to strengthen weak skeletal muscle contractions. An autoimmune disease called myasthenia gravis, in which there is reduced nicotinic receptor function, is treated with the AchE inhibitor neostigmine. This drug can also be used to reverse the effects of curare.
Excitation-contraction coupling
The process by which an action potential in skeletal muscle fibers induces contraction is called excitationcontraction coupling (Fig. 7.9). The action potential migrates along the Sarcolemma and down the T tubules. At the triad, the action potential triggers the release of Ca2+ from the terminal cisterns of the SR.
The release of Ca2+ from the terminal cisterns involves the direct mechanical connection between the T tubules and the terminal cisterns (also called lateral sacs) of the SR. Located on the T-tubule membrane is a T-tubule voltage sensor, which senses a change in membrane potential associated with the action potential. A change in voltage causes a conformational change in the T-tubule voltage sensor that is transmitted to the Ca2+ channels on the terminal cisterns of the SR, causing them to open and release Ca2+ into the cytosol. This direct mechanical connection is unique to skeletal muscle, whereas smooth muscle has a different mechanism and different source of calcium for excitati on-contraction coupling.
Cytosolic calcium levels increase at least 10-fold. As cytosolic calcium levels increase, Ca2+ binds to troponin, causing a conformational change in the shape of this globular protein. This change in shape allows tropomyosin to slide into the grooves of the double helix formed by F-actin (Fig. 7.10). As tropomyosin slides into the groove, it uncovers the myosin-binding sites on G actin. Once uncovered, the heads of the myosin filament binds to the myosin-binding sites, and contraction begins.
When stimulation from the motor neurons ends, the action potential is no longer propagated down the T tubules. At this point, a calcium-active transport pump called calsequestrin actively pumps Ca2+ back into the terminal cisterns of the SR. This process requires ATP, and allows for the concentration of Ca2+ in the SR to be 10,000 times higher than in the sarcoplasm. As the Ca2+ levels in the sarcoplasm decrease, troponin returns to its resting configuration, and tropomyosin again covers the myosin-binding sites on the G actin.
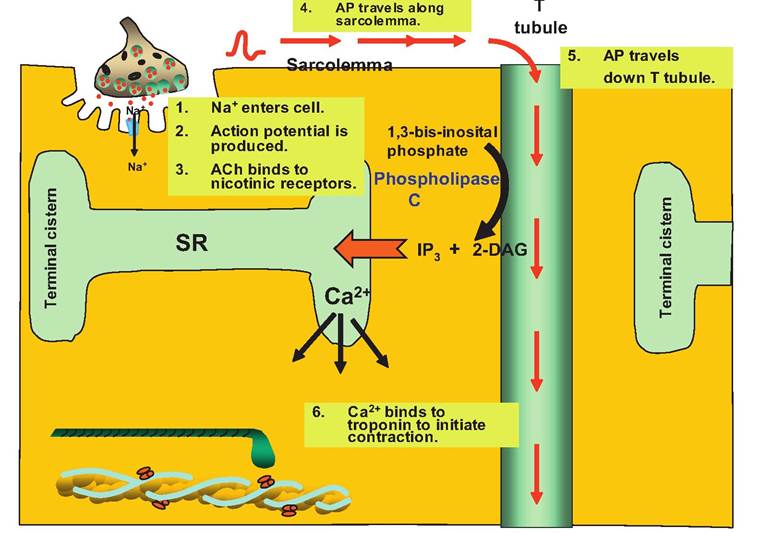
Fig. 7.9. Increasing sarcoplasmic calcium concentration. The action potential (AP) migrates along the Sarcolemma and down the T tubule. When reaching the triad, the AP activates the enzyme phospholipase C, resulting in the production of 2-diacylglycerol (2-DAG) and inosital triphosphate (IP3). The 2-DAG remains membrane bound while IP3 diffuses through the sarcoplasm to the terminal cistern of the SR. This opens Ca2+ release channels, causing the release of Ca2+ from the SR into the sarcoplasm. Ca2+ then binds to troponin, which initiates contraction.
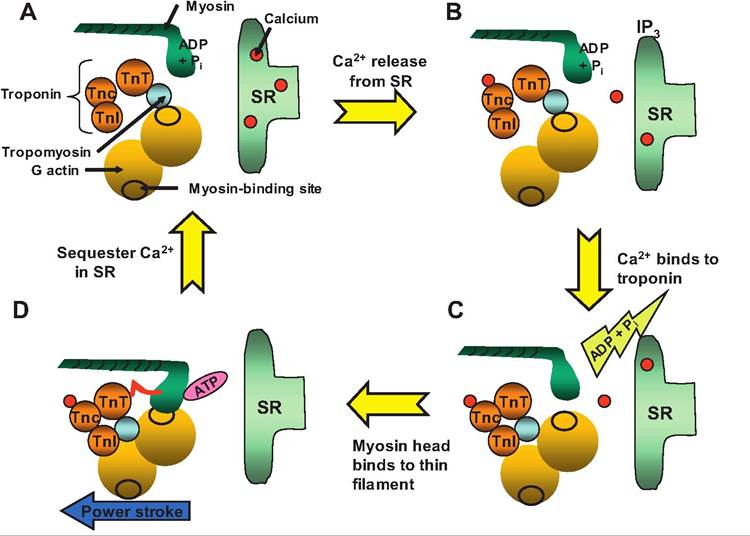
Fig. 7.10. Excitation-contraction coupling. (A) At rest, calcium is sequestered in the SR, and the myosin head sits perpendicular to the thin filament.
The myosin head is a charged intermediate with ADP and inorganic phosphate (Pi) attached. (B) Calcium released from the SR binds to the TnC component of troponin. (C) The conformational change in troponin results in the tropomyosin filament sliding into the groove of the double helix formed by F actin, thus uncovering the myosin-binding site located on the actin filament. The myosin head binds to actin, releasing ADP and Pi. (D) The power stroke occurs when the myosin head tilts toward the M-Iine.Sliding filament theory
During skeletal muscle contraction, the length of the thin and thick filaments does not change. Instead, the thin filaments slide between the thick filaments as the myosin heads "grab" the actin filaments and pull them toward the M-Iine (Fig. 7.11). Hence, as the thin filaments move toward the M-line, the Z-Iines get closer, thus decreasing the length of the sarcomere and the myofibril. As the sarcomere width decreases, the muscle shortens.
Contraction of skeletal muscle involves four steps:
1. Hydrolysis of ATP. In the resting position, the myosin head is perpendicular to the thin filament, and ATP has been hydrolyzed to ADP and inorganic phosphate (Pi), creating a charged intermediate. The myosin head is "waiting" for a binding site to become available on the thin filament.
2. Formation of cross bridges. When tropomyosin uncovers the myosin-binding sites on the thin filament, the myosin head binds to one of these sites liberating ADP and Pi.
3. Power stroke. Release of Pi initiates the power stroke in which the myosin head tilts toward the M-line, and ADP is released. The myosin head thus pulls the thin filament toward the M-Iine so that there is greater overlap between the thick and thin filaments. Hence the name, the sliding filament theory.
4. Detachment of the myosin head. At the end of the power stroke, the myosin head remains attached to actin until a molecule of ATP attaches to the myosin head, thus breaking the bond between myosin and actin.
The myosin head returns to its perpendicular position as it hydrolyzes ATP, thus returning to step 1.This cycle then repeats itself as long as the myosin- binding sites on the actin remain uncovered and sufficient ATP is present. Each thick filament possesses about 600 myosin heads. As contraction occurs, these heads are attaching and detaching throughout the cycle such that at any given time, there are many myosin heads attached. Therefore, contraction force is always being generated during this time. The myosin heads are sequentially "walking" the thin filament toward the M-Iine throughout the contraction cycle, and therefore pulling the Z-Iines closer together.
The elastic components in muscle include titin, tendons, and the connective tissue sheaths (endomysium, perimysium, and epimysium). As the muscle fibers contract, the elastic components are stretched. This stretch is relayed out to the tendons, which then pull on the bones causing them to move.
As calcium is sequestered in the SR, tropomyosin again covers the myosin-binding sites. Hence, the myosin head, with its hydrolyzed molecule of ATP attached, assumes its resting position poised to attach to actin when a binding site becomes available. As contraction ceases, the elastic components of the muscle help return the muscle to its resting position.
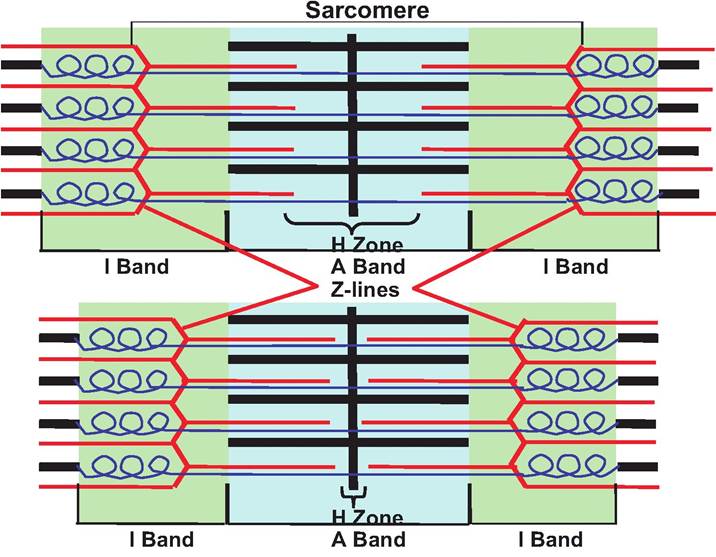
Fig. 7.11. Sliding filaments. The top sarcomere is at rest while the bottom sarcomere is in a contracted state. Note that in the contracted state, the Z-Iines move closer together and the I bands and H zone shorten while the A band remains the same width.
Rigor mortis
After death, the supply of energy within the cells diminishes as metabolism ceases. As such, the cells can no longer synthesize ATP. ATP is needed to actively remove sequestered Ca2+ into the terminal cisterns of the SR. In addition, since the cross bridges can be broken only in the presence of ATP, myosin and actin remain attached following death. This occurs in all the skeletal muscles, creating a state called rigor mortis (rigidity of death) in which the animal appears rigid. Rigor mortis ceases only as proteolytic lysosomal enzymes released by autolysis digest the cross bridges.
Summary of skeletal muscle contraction
The following steps summarize the contraction of skeletal muscles:
1. Release of ACh. Stimulation of the α-motor neuron to skeletal muscle causes release of ACh at the NMJ.
2. Activation of nicotinic receptors. ACh binds to nicotinic receptors causing an influx of Na+ into the muscle fiber resulting in the generation of an end-plate potential.
3. Generation of an action potential. Generation of an end-plate potential results in an AP developing in the muscle fiber.
4. Release of Ca2+ from the SR. The AP is propagated along the Sarcolemma and down the T tubules where it causes the production of IP3 at the triad. IP3 then diffuses to the SR where it causes the release of Ca2+ into the sarcoplasm.
5. Uncovering myosin-binding sites. Ca2+ binds to troponin causing a conformational change resulting in the movement of tropomyosin into the grooves of the thin filament, thus uncovering the myosin-binding sites on the thin filament.
6. Power stroke. Once uncovered, the heads of the myosin filaments bind to their binding sites on the actin filament. Once bound, the myosin head tilts toward the M-line, dragging the thin filaments in the same direction.
7. Breaking the myosin-thin filament bond. If present, ATP binds to the myosin head, which breaks the bond between myosin and the thin filament.
8. Termination of ACh activity. At the conclusion of motor neuron stimulation, ACh is broken down in the synaptic cleft by the enzyme AChE.
9. Sequester calcium. At the conclusion of motor neuron stimulation, calcium is actively sequestered back into the terminal cisterns of the SR by the protein calsequestrin.
10. Myosin head returns to resting position. After detaching from the thin filament, the myosin head hydrolyzes ATP and returns to a position perpendicular to the thin filament. ADP and Pi remain bound to the myosin head until it attaches to another myosin-binding site on a thin filament.
Length-tension relationships
The tension developed by a muscle fiber during contraction is dependent on the length of the sarcomere prior to contraction. At a sarcomere length of approximately 2.0-2.4 μm, or 90-110% of the resting sarcomere length, the overlap between the actin and myosin filaments is optimal, and the muscle can generate the maximum tension (Fig. 7.12). At these lengths, the maximum number of cross bridges can be formed between the myosin head and thin filament. As the muscle is either contracted or stretched, the number of cross bridges that can form decreases, and less tension is generated during contraction.
As the muscle fiber is stretched to approximately 170% of its resting length, the thick and thin filaments no longer overlap, and therefore, no tension can be generated. Conversely, as the length of the sarcomere becomes too short, the thick filaments are compressed against the Z-line, decreasing the number of cross bridges that can be produced.
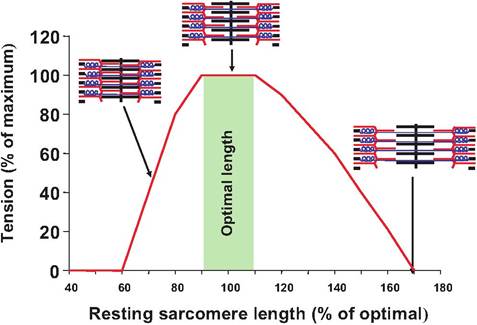
Fig. 7.12. Length-tension relationship. Attheoptimal resting sarcomere length, the maximum number of cross bridges can be formed between myosin and the thin filament resulting in the maximum tension. As the length of the sarcomere is stretched or compressed, the number of cross bridges is reduced, resulting in less tension.
A muscle twitch
Treppe
A single stimulation of a motor neuron results in a single contraction or twitch (Fig. 7.13). Although twitches can produce heat during shivering, they are not generally observed during normal muscle contraction. Instead, prolonged stimulation results in more tension being produced than caused by a single twitch.
A recording of a single muscle twitch is called a myogram. A single twitch can last from 20 to 200 ms, depending on the type of muscle, temperature, stretch of the muscle, and so on. A muscle twitch consists of three phases:
1. Latent phase. Following stimulation by the motor neuron, the action potential moves along the sar- colemma and down the T tubules to the triad where it induces the release of Ca2+ from the terminal cisterns of the SR. The calcium then diffuses to troponin where it binds. This process generally lasts about 2 ms.
2. Contraction phase. After a conformational change in troponin allows tropomyosin to move, thus uncovering the myosin-binding sites on the actin filament, the myosin head binds to the actin filament, and the power stroke pulls the thin filament toward the M-line, which produces tension. This lasts about 10-100 ms.
3. Relaxation phase. Calcium is sequestered into the SR, the tropomyosin covers the myosin-binding sites on the actin filament, and ATP causes the myosin head to detach from the actin filament, decreasing the number of cross bridges and tension. This phase can also last 10-100 ms.
If skeletal muscle is stimulated a second time, shortly after the relaxation phase of the first twitch, the second twitch will generate greater tension (Fig. 7.14). This increase in tension is known as treppe, German for "stairs." The increase in tension caused by subsequent stimulations results from the gradual increase in sarcoplasmic calcium since the calcium pumps located in the SR are unable to sequester all the calcium between twitches.
Summation
While a muscle twitch is an all-or-none response, muscle contraction is graded, meaning that it displays varying length and strength of contraction. There are two mechanisms leading to graded responses: (1) changing the frequency of stimulation and (2) changing the strength of the stimulus.
Wave summation
While muscle twitches can be observed in the laboratory, muscle contraction generally involves smooth sustained contraction resulting from frequent stimulation. If a second stimulation occurs before the muscle completes its relaxation phase, the second twitch will create greater tension than the original twitch. This process is called wave summation (Fig. 7.15). This generally occurs at stimulation rates of about 50 per second. Stimulation occurs rapidly enough that the SR
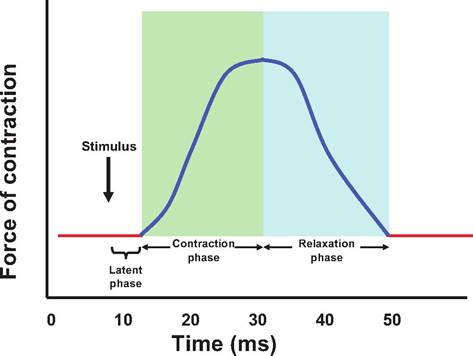
Fig. 7.13. Muscle twitch. A myogram showing the three stages of an isometric muscle twitch. The stimulus is followed by a latent period during which calcium is released from the sarcoplasmic reticulum (SR) and then binds to troponin. The contraction period is when the myosin head actively binds and pulls on the thin filaments, and the relaxation period occurs when calcium is sequestered in the SR.
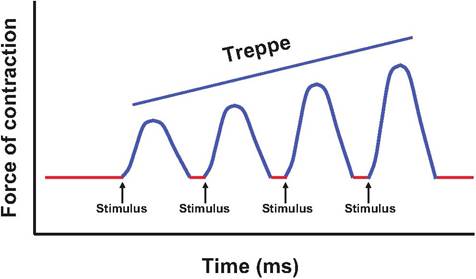
Fig. 7.14. Treppe. If the muscle is stimulated shortly after the relaxation phase, the subsequent muscle twitches generate greater tension, producing a steplike increase in magnitude called treppe. The increase in tension in subsequent twitches results from the increase in sarcoplasmic calcium due to the inability of the SR to recapture all the calcium between twitches.
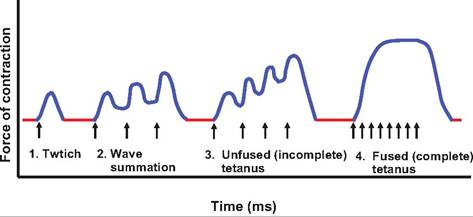
Fig. 7.15. Wave summation and tetanus. (1) A single twitch.
(2) The muscle is stimulated (T) before the relaxation phase is complete, causing wave summation and increased contraction force. (3) Frequency of stimulation is more rapid, resulting in unfused tetanus in which the individual twitches can still be discerned. (4) Frequency Ofstimulation is so rapid that individual twitches cannot be distinguished, resulting in fused tetanus.
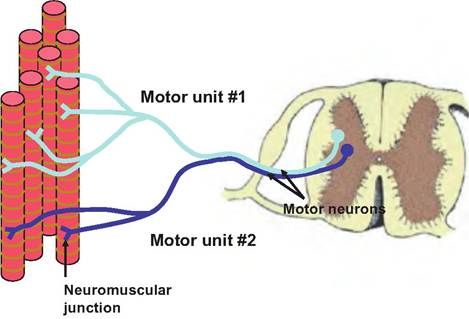
Fig. 7.16. Motor units. All of the muscle fibers innervated by a single motor neuron constitute a motor unit. Motor unit #1 is larger than motor unit #2 because it innervates more muscle fibers. Motor unit #2 would be involved in more precise motor movement than motor unit #1.
is no longer able to sequester Ca2+ between twitches. In addition to prolonging contraction, the second contraction causes greater shortening than the first contraction because it is superimposed on an already contracted muscle, thus increasing tension.
As the frequency of stimulation increases, the tension developed also increases. Incomplete tetanus (tetan = rigid or tense) occurs when the individual twitches are still distinguishable. When the frequency of stimulation is rapid enough to eliminate the relaxation phase, and the individual twitches are no longer distinguishable, the contraction is termed complete tetanus. Complete tetanus is the normal state observed during muscle contraction. Wave summation results in smooth, continuous muscle contraction. Note that the frequency of nerve stimulation cannot be faster than the absolute refractory period of the neurons.
Multiple motor unit summation
A second type of summation that increases the force of muscle contraction is called multiple motor unit summation or recruitment. Skeletal muscles have thousands of muscle fibers. All of the muscle fibers innervated by a single motor neuron constitute a motor unit (Fig. 7.16). The size of a motor unit can vary with a single motor neuron innervating as few as four to six muscle fibers, or as many as several thousand. Where fine, delicate movements are necessary, such as in the lips, motor units are small, whereas in areas where precise movements are not necessary, such as in the hindquarter of a beef cow, the motor units are much larger. Motor units are intermingled within a muscle so that they always deliver force on the tendon attaching to a bone.
As an animal begins a task, it generally stimulates the smallest motor units. However, if more force is required, more and larger motor units are recruited in order to increase the tension produced by the muscle.
Muscle tone
Skeletal muscle is seldom flaccid, but instead maintains a degree of tension called muscle tone. Since skeletal muscle contraction is controlled by motor neurons releasing ACh, muscle tone is established by the central nervous system. If these motor neurons are cut, skeletal muscle becomes flaccid. Muscle tone is due to the alternating stimulation of motor units by the central nervous system. Such tone helps keep an animal upright, keep the head held up, stabilize joints, and maintain posture.
Muscle tone is not unique to skeletal muscle, but it is also found in smooth muscle. For example, blood vessels generally maintain a vascular tone as does the gastrointestinal tract.
Isometric versus isotonic contraction
There are two major categories of muscle contraction: isotonic and isometric (Fig. 7.17). During isotonic (iso = same; tonos = tension), the length of the muscle changes as force is generated, resulting in movement. There are two types of isotonic contractions, concentric and eccentric. In a concentric contraction, the muscle gets shorter as it works. In other words, the muscle forms cross bridges and the thin filaments interdigitate within the thick filaments, overcoming
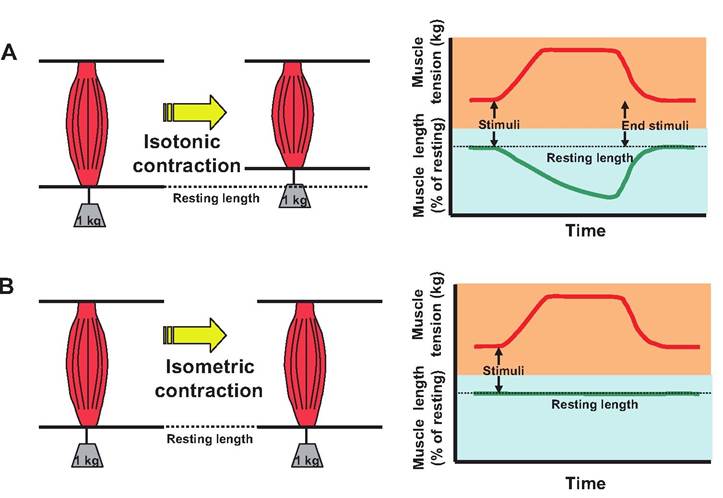
Fig. 7.17. Isotonic versus isometric contractions. (A) During isotonic contraction, force is generated as the muscle shortens. (B) In isometric contraction, force is generated, but there is minimal shortening of the muscle.
the resistance of the load on the muscle. During eccentric contraction, the tension developed by the muscle is less than the load on the muscle. As a result, the muscle lengthens. As an animal walks down a steep incline, the animal controls the rate of elongation of muscles as the legs stretch to the next location.
During isometric (metric = measure) contraction, the length of the muscle does not change because the tension produced does not exceed the resistance. Isometric contraction is commonly observed in postural muscles that maintain a constant body position while opposing gravity.
When performing various movements, animals use all these types of contractions. Consider the motions as a dog sits and then stands back up. The quadriceps are involved in controlling this motion. As a dog begins to sit, the knees begin to bend or flex (eccentric). As the dog holds a position partway through the sitting motion, an isometric condition exists. As the dog stands, thus extending the knee, isometric and concentric contractions occur.
Muscle relaxation or return to resting length
While muscle contraction is an active process requiring energy, the relaxation of muscle is a passive process. Elastic forces, opposing muscles, and gravity act to return the muscle to its resting length. Such elastic fibers include connective tissue and many of the muscle proteins, such as titin.
Contraction of the opposing muscle will also help return a muscle to its resting length. For example, as the triceps brachii muscle in the back of the front leg contracts, it causes the biceps brachii on the anterior portion of the leg to extend.
Similarly, gravity can cause muscles to extend. The neck of a horse is extended in order to look upward, and then when the muscles are relaxed, the head will move toward the ground, thus stretching the neck muscles that originally were involved in extension.