Skeletal muscle
Skeletal muscle accounts for approximately 40% of body weight. Each skeletal muscle is considered an organ and is made up of muscle fibers as well as connective tissue, blood vessels, and nerve fibers.
Connective tissues
As with neurons, each muscle has three connective tissue layers (Fig. 7.1).
1. Epimysium. The entire muscle is surrounded by a dense, irregular connective tissue layer called the epimysium containing a dense concentration of collagen fibers. This layer separates the muscle from surrounding tissue.
2. Perimysium. In cross section, a muscle consists of multiple groupings of muscle fibers called fascicles (bundles). Each fascicle is surrounded by the perimysium (peri = around) containing collagen and elastic fibers. This layer contains blood vessels and nerves supplying the fascicles.
3. Endomysium. Within each fascicle are individual skeletal muscle cells, called muscle fibers, each surrounded by the endomysium (endo = within). Within this connective tissue layer are capillaries
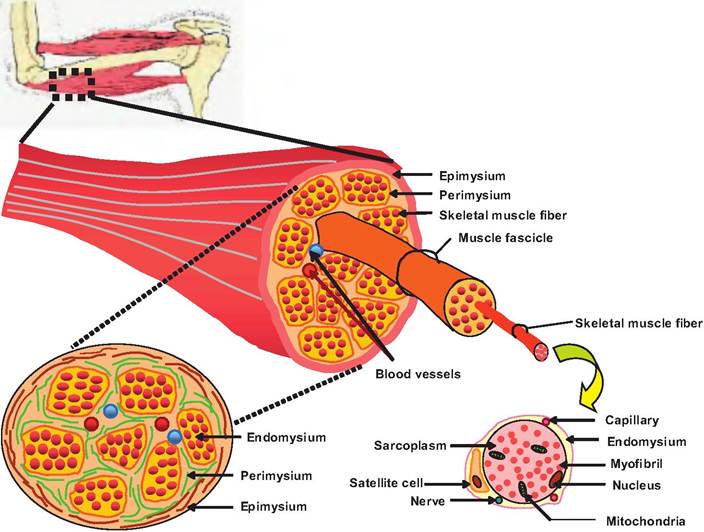
Fig. 7.1. Connective tissue sheaths in skeletal muscle. Each skeletal muscle represents skeletal muscle fibers grouped into a muscle surrounded by a thin connective tissue sheath called the epimysium. Within the muscle are groupings of muscle fibers called fascicles, which are surrounded by the perimysium. Within each fascicle are individual muscle fibers surrounded by the endomysium.
supplying each muscle fiber, nerve fibers controlling the muscle, and satellite cells. These latter cells serve as stem cells that can help repair damaged muscle.
Near the ends of the muscle, the epimysium, perimysium, and endomysium blend together forming either a bundle called a tendon or a broad sheet called an aponeurosis. Tendons and aponeuroses attach muscle to bones, blending with the periosteum of the bone.
These attachments allow contraction of the muscles to move the bones.Blood vessels and nerves
The two innermost layers of connective tissue within the muscle each contain blood vessels and nerves. Skeletal muscle is generally under voluntary nervous control, and therefore requires stimulation for nerve fibers to initiate contraction. Therefore, individual nerve fibers must innervate each muscle fiber to control contraction. While the diaphragm consists of skeletal muscle, it usually is under involuntary control but can be under voluntary control as well.
Skeletal muscle fibers
Skeletal muscle tissue consists of large, multinucleated cells commonly referred to as muscle fibers (Fig. 7.2).
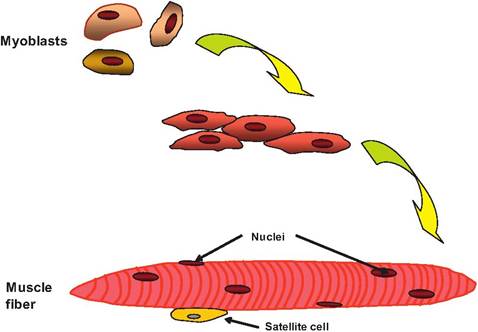
Fig. 7.2. Formation of skeletal muscle cell. During embryonic development, myoblasts begin to fuse forming a large, muItinucleated skeletal muscle cell called a muscle fiber. Nonfused myoblasts remain as satellite cells that function as muscle stem cells.
Muscle fibers can be 100 μm in diameter and run the entire length of a muscle, and can contain hundreds of nuclei. These cells form from the fusion of small, individual muscle cells called myoblasts during development. Instead, some remain unfused and become satellite cells. While skeletal muscle fibers are incapable of dividing, new muscle fibers are produced from satellite cells located in the adult muscle. Not all myoblasts fuse to form muscle fibers. The satellite cells can later enlarge, divide, and then fuse with damaged muscle cells, thus regenerating the muscle.
The nuclei are located immediately under the plasma membrane, which in skeletal muscle is called the Sarcolemma. There is a resting membrane potential present due to the unequal distribution of ions across the Sarcolemma similar to that found in neurons. The cytoplasm of skeletal muscle is called sarcoplasm. Within the sarcoplasm are glycosomes, storage granules of glycogen, and myoglobin, a red pigmented protein that stores oxygen.
Although skeletal muscle fibers are large, an electrical signal must be able to propagate throughout the cell quickly to cause contraction. Transverse, or T, tubules are small-diameter tubes running perpendicular to the Sarcolemma and traveling into the sarcoplasm. These tubes are Continuouswiththe extracellular space, and thus they contain extracellular fluid. They can be thought of as extensions of the Sarcolemma. As we will see later, the action potential can travel along the Sarcolemma and down the T tubules.
Myofibrils
Muscle fibers are composed of functional subunits called myofibrils. Each muscle fiber contains hundreds to thousands of myofibrils that run longitudinally the length of the fiber. The myofibrils consist of bundles of myofilaments that are protein filaments composed primarily of actin and myosin, the two contractile proteins in muscle. Actin forms the bulk of the thin filaments, and myosin forms the bulk of the thick filaments. The myofibrils are packed tightly into the muscle fiber, forcing the mitochondria, nuclei, and other organelles to be squeezed toward the outer edge of the cell.
The myofibrils contain three types of proteins that will be discussed in more detail when describing the structure of thin and thick filaments below:
1. Contractile proteins. Contractile proteins generate the force during contraction. These proteins include myosin and actin.
2. Regulatory protein. Regulatory proteins help initiate and terminate the contraction process and include tropomyosin and troponin found on the thin filaments.
3. Structural proteins. Structural proteins help maintain the alignment of the thin and thick filaments, provide elasticity and extensibility, and attach the myofibrils to the sarcolemma. These proteins include titin, myomesin, and dystrophin.
The myofibrils are attached to the inner surface of the sarcolemma. The outer surface of the muscle fibers is attached to collagen fibers that help connect the cells to the tendon or aponeuroses.
Therefore, as the muscle fibers contract, they exert force on the bones causing them to move.Sarcoplasmic reticulum
Similar to the endoplasmic reticulum in nonmuscle cells, the sarcoplasmic reticulum (SR) forms a tubular network surrounding each myofibril (Fig. 7.3). The terminal cisternae (end sacs) of the SR are always found in pairs, with an intervening T tubule. The combination of a terminal cisterna, a T tubule, and the adjacent terminal cisterna form a triad. Note that the T tubule communicates with the extracellular space while the SR is intracellular.
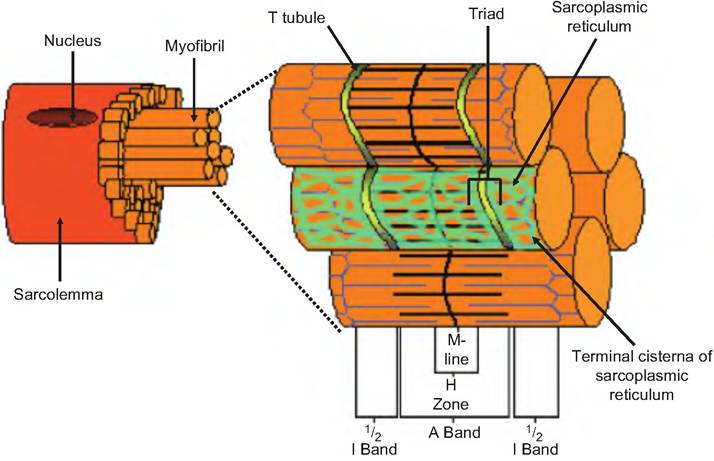
Fig. 7.3. T tubule and sarcoplasmic reticulum. The transverse, or T tubules are inwardly directed invaginations of the sarcolemma found near the junction of the A and I bands. The sarcoplasmic reticulum is a network of tubules found inside the cell and which have terminal cisternae near the T tubules. Two terminal cisterna and the intervening T tubule make a triad.
The terminal cisternae have an active calcium pump that pumps calcium from the sarcoplasm into the SR. This maintains a low concentration of free calcium within the sarcoplasm, whereas the free calcium concentration inside the SR may be 1000 times greater. Also found within the terminal cisternae is the protein calsequestrin that reversibly binds Ca2+. The free and calsequestrin-bound calcium concentrations can be 40,000 times that in the sarcoplasm. As we will discuss later, the terminal cisternae of the SR are the source of calcium for skeletal muscle contraction.
Sarcomeres
The functional unit of skeletal muscle is the sarcomere. A myofibril consists of thousands of sarcomeres (Fig. 7.4). In stained cross sections of skeletal muscle, alternating light and dark bands are evident, which are called the I band and A band, respectively. These bands give skeletal muscle its striated appearance. The dark bands alter the plane of the polarized light and are therefore anisotropic (i.e., not having the same properties in all directions), whereas the light bands do not alter the plane of polarized light and are therefore isotropic (i.e., appear the same in all directons), thus the names A band and I band, respectively.
The sarcomere is composed of thick and thin filaments, proteins that stabilize those filaments, and proteins that regulate the interactions between thick and thin filaments. As shown in Figure 7.4, a sarcomere is the region between two adjacent Z discs (or Z-Iines). It consists of one-half an I band, an A band, and one- half an I band. The A band is the length of the thick filament, and can contain both thick and thin filaments. In a muscle at rest, a lighter region can be found in the center of the A band called the H zone (from 1ιelle, meaning bright), which contains only myosin. This region disappears as skeletal muscle contracts and the actin filaments overlap, thus entering this area. The M-line, named for being in the middle of the sarcomere, transects the H zone and is composed of proteins that stabilize the position of the thick filaments. Near the ends of the A band are zones of overlap where thin and thick filaments are found side by side.
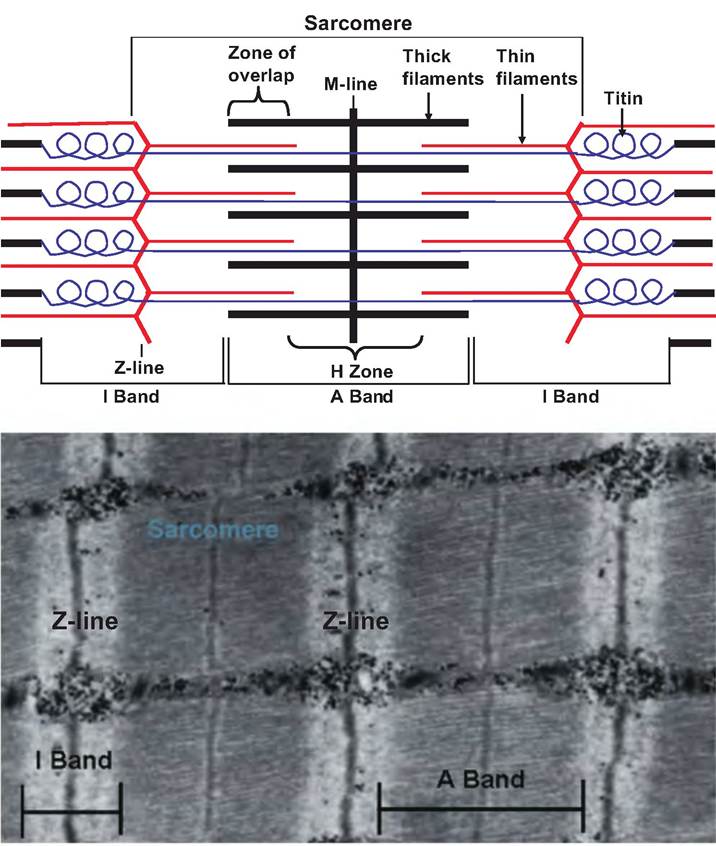
Fig. 7.4. Sarcomere. A sarcomere, the functional unit of skeletal muscle, runs from Z-Iine to Z-Iine. The various segments of the sarcomere are identified in the top portion of the figure, and a photomicrograph of a sarcomere is shown in the bottom portion of the figure.
The I band, located between each intervening A band, contains thin filaments. The I band is bisected by the Z-Iine that consists of proteins called actinins, which interconnect adjacent thin filaments.
There are several structural proteins associated with the myofibrils making the sarcomere. Titin (from titan, meaning gigantic) is a large protein, and the third most abundant protein in the sarcomere behind myosin and actin. Each titin molecule extends from the Z-Iine to the M-Iine and helps anchor a thick filament to both the Z-Iine and M-line. This provides stabilization for the position of the thick filaments. As shown in Figure 7.4, the portion of the titin molecule located between the Z-Iine and the end of the thick filament is very elastic and can stretch up to four times its resting length.
Therefore, titin probably assists in returning the muscle to its resting length following stretching.The Z-Iine is composed of the protein nebulin. Nebulin anchors thin filaments and connects myofibrils to each other throughout the muscle cell. The M-Iine is composed of the protein myomesin. The M-Iine binds to titin, thus helping to connect adjacent thick filaments. Dystophin is another structural protein that links thin filaments to integral membrane proteins in the Sarcolemma. Other proteins in the sarco- Iemma then attach to the connective tissue sheath surrounding the muscle. Thus, the contractive forces generated in the sarcomere are transferred throughout the muscle.
Thin filaments
Thin filaments are 5-6 nm in diameter and 1 μm in length (Fig. 7.5). Each thin filament is composed of four proteins:
1. F actin. Thin filaments are composed of two strands of F actin, also called filamentous actin, arranged in a double-stranded helix. Each strand
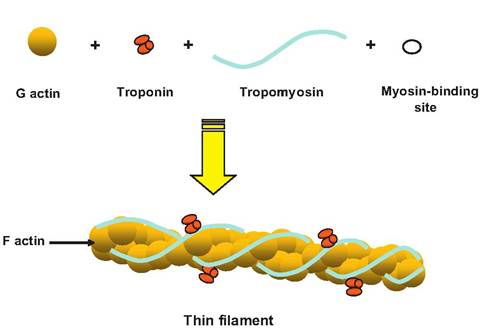
Fig. 7.5. Thin filament. The thin filaments in skeletal muscle consist of G actin, troponin, and tropomyosin. G-actin polymerizes into F actin, or filamentous actin. Troponin is made of three globular proteins binding G actin, tropomyosin, and calcium ions, respectively. Two strands of tropomyosin, a rod-shaped protein, intertwine around the F actin covering the myosin-binding sites while the muscle is at rest.
of F actin is composed of polymers of G actin, or globular actin. Therefore, the F actin appears as two twisted strands of pearls, with each pearl being analogous to a molecule of G actin.
2. Tropomyosin. Strands of tropomyosin (trope = turning) wrap around the length of the F actin. Each tropomyosin molecule is a double-stranded protein that, at rest, covers seven myosin-binding sites on the actin filament.
3. Troponin. A globular protein, troponin consists of three subunits. One binds to tropomyosin (TnT), one to G actin (TnI), and the other to calcium ions (TnC). Therefore, troponin controls the structural relationship between tropomyosin and F actin. At rest, troponin allows tropomyosin to be positioned such that it covers the myosin-binding sites. When a muscle is stimulated and intracellular calcium levels increase, calcium binds to troponin, causing a conformational change that allows the tropomyosin to slide into the grooves of the double helix of actin and thus uncover the myosin-binding sites.
Thick filaments
Thick filaments are 10-12 nm in diameter and 1.6 μm in length (Fig. 7.6). Thick filaments consist of approximately 500 myosin molecules, each composed of two myosin subunits wrapped around each other. The long tails of the myosin molecules line up forming the thick filament, and the heads of the myosin molecules project off the filament toward the adjacent thin filaments. The head of the myosin molecule consists of two globular proteins, has ATPase activity, and is able to bind to the actin filament. A cross bridge is formed
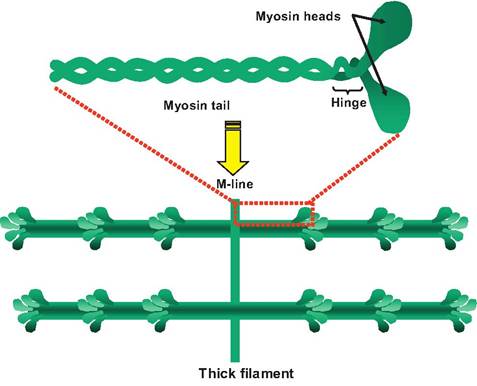
Fig. 7.6. Thick filament. A single myosin molecule is shown at the top. It contains a pair of intertwined subunits each consisting of a tail, a hinge region, and a globular head. The thick filaments contain approximately 500 myosin molecules in which the tails are lined up so that the heads project away from the M-Iine.
when the head of the myosin binds to the actin filament. There is a hinge between the head and the tail of the myosin molecule that allows the head to pivot toward or away from the M-Iine.
The myosin molecules are arranged so that their tails point toward the M-Iine. In the H zone, there are no myosin heads, only tails. Also within each thick filament is a molecule of titin extending from the M-Iine to the Z-Iine.
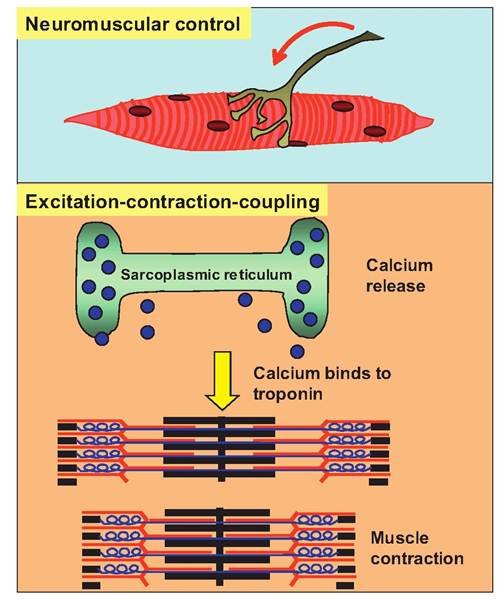
Fig. 7.7. Summary of skeletal muscle contraction. Stimulation of α-motor neurons going to skeletal muscle causes the release of acetylcholine at the neuromuscular junction. This causes the production of an action potential in the muscle fiber that spreads along the Sarcolemma and down the T tubules where it causes the release of calcium ions from the sarcoplasmic reticulum. Calcium then diffuses to the thin filaments where it binds to troponin to initiate contraction.