MiIk synthesis and secretion
The mammary gland is an unusual exocrine gland in several respects. The product is a very complex mixture that depends on apocrine and meocrine modes of secretion. Other components are derived by passage of soluble molecules across (transcellu- lar) and sometimes between (paracellular) the cells.
Physically, milk is a complex solution of salts, carbohydrates, miscellaneous compounds with dispersed proteins and protein aggregates, casein micelles, and fat globules. Milk osmolarity generally equals blood (~300 mθsm) and has a pH between 6.2 and 7.0. Bovine and human milk have an average pH of 6.6 and 7.0, respectively.Once initiated, milk secretion continues more or less continuously throughout lactation. This secretion mixture includes membrane-bound lipid droplets; casein micelles; and an aqueous phase usually containing lactose, minerals, other proteins, and a variety of other soluble components. Milk is stored within the lumen of the alveoli and ductular system until it is removed by the milking machine or the suckling offspring. Suckling intervals vary widely between mammals, ranging from minutes to hours in cattle, to once daily in rabbits, to once every 2 days in tree shrews, or to only once a week in some seals. Moreover, although there are species-specific changes in milk composition with stage of lactation, milk composition for most of lactation is generally only moderately affected by environmental or nutritional changes, despite the often-dramatic changes in milk volume. Function of the mammary gland during established lactation is closely linked with a number of hormones,
Adapted from Jenness (1974) and Oftendal (1984, 1997).
growth factors, and local tissue regulators, but it is difficult to ascribe a specific transport activity to a particular molecule or to determine if effects are direct or indirect.
Across species, there are dramatic differences in milk composition. Milk from Holstein cows (the source of the majority of milk for human consumption in Western societies) has about 3.2% protein, 3.4% fat, and 4.6% lactose. In contrast, hooded seals produce milk with about 6.0% protein, 50% fat, and virtually no carbohydrate. Table 18.7 provides milk composition data for some selected species.
Along with mammary cell-specific constituents, milk contains a myriad of minor components. Many of these molecules are important nutrients or regulators of the neonate (growth factors, water, and ions), but other components may include drugs or other Xenobiotic substances transported from the circulation. Molecules are transported into the milk by several possible routes. Mammary epithelial cells are able to maintain substantial gradients for Na+, K+, and Cl ions across the cell membrane. During established lactation there are also gradients between milk and plasma. These ions are important to maintain normal electrical gradients across the alveolar cell membranes, but they also are critical in regulation of milk osmolarity, especially for those species with low lactose production. Concentrations of Na+inside (~43mM) the cells are typically lower than outside (150 mM), but the gradient for K+ is the opposite (143 mM inside compared with 4.5mM outside). These differentials are maintained by action of Na+ K+ ATPase pumps in the basolateral membranes. The apical plasma membrane is permeable to both ions so that the distribution of these ions into milk is controlled by the electrical potential across the apical plasma membrane. Milk is electrically positive with respect to the cell, so that the concentrations of Na+ and K+ are lower in milk than in the cells, but the K+∕Na+ ratio (-3:1) is similar. Concentration of CP is higher inside the cells than the equilibrium distribution would suggest, so membrane pumps in the basolateral and apical membranes act to sequester CP.
It is easy to imagine that this balance of ions between milk and blood is readily compromised if the leakiness of the tight junctions is altered (Stel- wagen, 2001).As dietitians and nutritionists frequently note, milk is a rich source of calcium, with total concentrations equaling IOOmM or more. With the onset of lactation the mammary gland extracts large quantities of calcium to supply the developing neonate. For high- producing dairy cows, this demand can lead to metabolic periparturient paresis unless animals are carefully managed. The calcium in the milk exists as free calcium, casein-bound calcium, or calcium associated with various inorganic anions, for example, citrate and phosphate. There is little movement of calcium from milk to blood, which suggests that calcium cannot pass across the apical plasma membrane. Given that most of the calcium is associated with the casein micelles, the Golgi vesicle route of secretion is the predominating pathway. However, since all of the milk calcium is derived from the circulation, there must be differences in transport between basolateral and apical membranes. Mammary cells maintain a low intracellular free calcium concentration in spite of the marked accumulation of calcium in milk. This is Importantbecause changes in free calcium concentration are closely linked with several hormone and growth factor-signaling pathways. One idea is that the rate of calcium influx into the cell is matched by a corresponding uptake of calcium by cellular organelles. The presence of an ATP-dependent calcium pump on Golgi membranes has been demonstrated. Calcium uptake by mammary cells is also likely hormonally regulated since parathyroid hormone-related protein and 1,25-(OH)2 vitamin D3 stimulates the uptake of calcium in cultured mammary tissue.
Lactation curves
Once initiated, lactation depends on regular suckling or milking of the mammary gland to maintain the lactation. Although the time required for regression varies markedly between species, that is, days for rodents versus weeks for ruminants, without milk removal the alveolar structure is eventually degraded, alveolar cells de-differentiate and many cells undergo apoptosis.
Without the stimulus of another gestation, the gland progressively reverts to a structure similar to that of the mature virgin. However, in dairy cows, milk production increases with each successive lactation milk. This suggests accumulative mammary growth with each lactation cycle. Normal husbandry dictates that dairy cows are rebred soon after the onset of lactation and milked for much of the concurrent pregnancy. Consequently, during the later part of lactation, the cow has dual functions of growth of the developing calf and continued lactation. Compared with wild ruminants or beef cows, the period between consecutive lactations is relatively short. Consequently, there is less opportunity for mammary regression or involution. Second, hormonal and metabolic changes associated with late gestation and preparation for parturition are conducive for mammary development. This means that the time course of mammary involution is impacted by concurrent gestation.Although early sections provide an overview of mammary development, the somewhat unique anatomy of the udder deserves special attention. In the cow and other ruminants the mammary glands are clustered together into groups of two (goats or sheep) or four (cattle) mammary glands to create the udder. This arrangement provides a practical advantage. Since the mammary glands and teats are close together, the portion of the milking machine attached to the animal (teat cups and teat cluster) can be relatively compact. For those not familiar with milking and management of modern dairy cows, the udder of a lactating Holstein cow can be massive. It is not unusual for a single cow to yield 30 kg or more of milk at a single milking. Combined with the mass of the udder tissues this means that the connective tissues of the mammary glands have to support as much as 70 kg of tissue and stored milk just before milking. Given the dorsal inguinal orientation of the udder, this is no trivial matter. Support is provided by strong flat suspensory ligaments, which are attached to the pelvic bone and to the strong tendons of the abdominal muscles in the pelvic area.
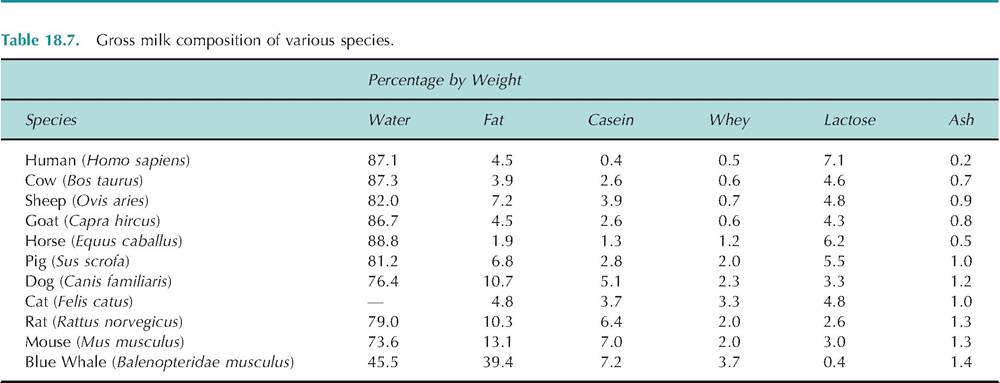
The udder is divided into two distinct halves, separated by the medial or median suspensory ligament, which provides most of the strength to hold the udder attached to the dorsal body wall. Fibers of the lateral ligaments are continuous with the medial ligament but spread over either side of the udder so that it appears to be held in a sling of connective tissue. The medial ligament is somewhat elastic but the lateral ligaments are not. As the milk accumulates in the udder, the normally vertical orientation of the teats is lost as teats progressively protrude laterally. As animals age, excessive degradation of the fibers of the medial suspensory ligament can reduce its support capacity so that the udder becomes pendulous irrespective of time relative to milking. This can lead to difficulties with milking, that is, problems maintaining attachment of teat cups as well as problems with teat injury and increased mastitis risk. Figure 18.12 illustrates the appearance and strength of the medial and lateral ligaments. The mammary glands of the udder are directly connected to the abdominal cavity only via passage through the inguinal canals. These are paired narrow oblique passages through the abdominal wall on either side of the midline, just above the udder. These canals allow passage of blood and lymph vessels and nerves to the udder.
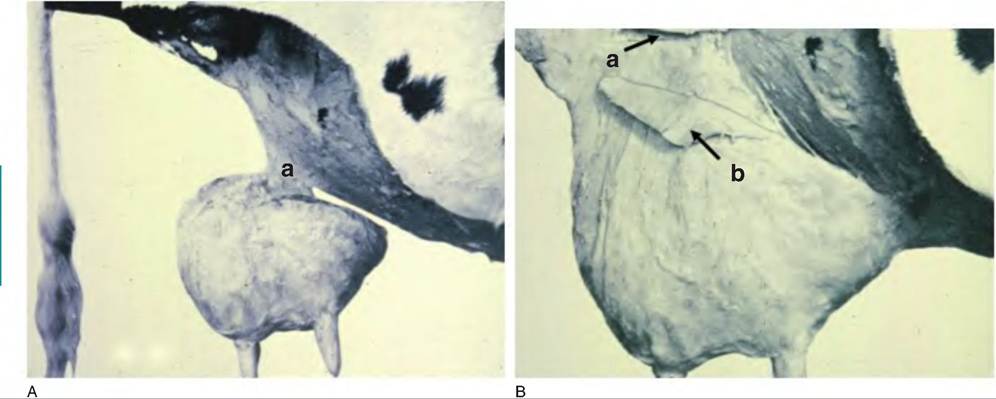
Fig. 18.12. Medial and lateral suspensory ligaments. Panel A shows the dissected udder of a cow supported by only the medial suspensory ligament (A). Panel B shows a portion of the lateral suspensory ligament dissected as a flap of tissue (arrows). The letters a and b indicate the cut edge and boundaries of a flap of the lateral suspensory ligament. Adapted from Swett et al. (1942).
Compared with most other mammary glands, those of animals with udders also have relatively large nipples or teats. Specifically, the teat of the ruminant has a single opening called the streak canal that leads directly into a space within the teat called the teat cistern.
The structure of the streak canal is considered further with respect to its role as the primary defensive barrier against mastitis. The space of the teat cistern would typically hold only a few milliliters of milk. Near the base of the teat there is an annular fold of tissue, which separates the teat cistern from the gland cistern. The gland cistern is roughly the size of an orange and holds ~200mL of milk. The gland cistern has many irregularly shaped cavities, which accommodate the endings of large intra-lobular ducts, which drain milk from the secretory tissue. Except for the terminal ducts that are directly adjacent to the alveoli, ducts are lined with at least two layers of nonsecretory epithelial cells. It is estimated that 40-60% of the milk is stored in the Iumenal spaces of the ducts or cisterns. Above the gland cistern, the tissue is progressively more dense and compact because of the relative lack of very large ducts and the closely arranged lobules.It has long been assumed that milk is prevented from escaping from the teat by the action of bands of sphincter-like smooth muscle cells in the teat meatus surrounding the streak canal. However, recent studies suggest that most of these smooth muscle cell elements are located some distance from the streak canal and that they are more likely involved in rhythmic contractions of the teat. Thus, closure of the teat canal depends on passive elastic elements in the region surrounding the streak canal. A more recent suggestion is for a multi-spiraled, net-like combination of elastic fibers and associated smooth muscle cells that produce a spiraling of the internal epithelial folds of the streak canal to affect closure. Regardless of the exact mechanism, the internal structure of the streak canal and its surface secretions prevent milk leakage and act as a barrier. Although milking ease and milk speed are important factors in the economics of a dairy operation, selection of cows with wide, short teat canals may well speed the milking process at the expense of increased risk of mastitis.
The skin of the teat is hairless but tough and resistant to tears or punctures. Histologically, it is a stratified squamous epithelium that extends over the teat end and into the teat opening for the length of the streak canal. However, relative lack of insulation seems to make the teats susceptible to cold weather problems. The washing of udders and teats in preparation for milking and movement of cows into freezing temperatures and winds before drying likely amplify teat injury problems. Between the outside skin of the teat and internal surface of the streak canal or teat cistern, the stromal tissue contains a network of blood vessels, lymphatic vessels, smooth muscle cells, and nerves. This extensive blood supply is important in maintenance of normal tissue temperature and especially so during severely cold temperatures. Because mechanical milking can retard blood flow in the teat and produce vascular congestion or swelling of the stromal tissue, this likely exacerbates problems with teat skin injury in cold weather.
The streak canal ranges from 7 to 16 mm in length with a mean diameter of 0.82mm (Fig. 18.13). The lowest 2 cm of the streak canal is especially important because of capacity of these tissues to act as a barrier to minimize milk leakage or entrance of environmental agents. Intuitively, the diameter of the streak canal is positively related to the rate of milk flow, but cows with the best balance of acceptable rates of milk flow and protection from bacterial invasion can expect greatest longevity in the herd. Teats vary in shape and size, from cylindrical to funnel shaped. Teat ends also vary, with some that are flat, but others round or pointed. Pointed teats are less common and are associated with slow milking times but resistance to mastitis. Round teats are common and occur on cows with faster milking times, but these cows exhibit some resistance to mastitis. Flat teat ends are less common, but these cows also tend to have faster milking with
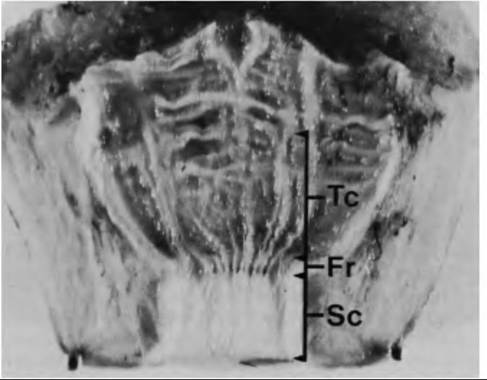
Fig. 18.13. Internal structure of the cow's teat. Areas illustrated include the streak canal (SC), Furstenberg's rosette (Fr), and the teat cistern (Tc). Adapted from Nickerson and Pankey (1983).
less resistance to mastitis. Certainly, the uniformity of milking machine teat cups means that cows with teats of large diameter or length are more likely to suffer milking machine-related trauma.
A lactation curve for typical Holstein cow is illustrated in Figure 18.14A. However, it is important to realize that estimates of milk yield need to be evaluated in relationship to milk composition, state of maturity of the offspring, suckling patterns, diet, and behavior of the species. Practically, production curves are most reliable for dairy animals (cows, goats, sheep) because these animals have been selected for ease of milking and handling, high yields of milk, and for their capacity to response to machine or hand milking. Because of their usefulness as bioreactors to produce pharmaceutical proteins in their milk, renewed interest in milking of pigs has emerged in recent years. Garst et al. (1999a, b) reported results for animals that were milked four times per day as well as effects on litter weight gains. Milk yields peaked at about 19 days postpartum and declined to 45 days postpartum. As might be expected, sows that were milked four times per day had litters with substantially lower weights at the time of weaning. Average daily milk yield per sow was 1.1 L per day or 43.1 ± 8.6L for the entire lactation period. There were also substantial differences in milk yields between individual mammary glands. For example, anterior glands produced more than posterior glands (-40 vs. 15 mL per milking). Examples of pig milk yields are illustrated in Figure 18.14.
For other species, direct measurement of milk yield requires that the animals be sedated and/or injected with exogenous oxytocin. Under these circumstances, the relevance of the data (other than comparison between treatments) can be questioned. Certainly, it is unknown if yields reflect "natural" levels of production or if measures of composition are accurate. An alternate for estimating yields is the response to timed suckling episodes. This typically involves removal of the suckling young for a period of time, then weighing the litter or offspring immediately before and after a set period when returned to the mother (Beal et al., 1990). For species in which the offspring are solely dependent on milk for nourishment, measurement of offspring weight gain is used as an indirect measure of milk production of the mother.
Fig. 18.14. Lactation curves in cows and sows. Panel A shows the lactation curve for a typical Holstein cow. Milk production typically peaks at about 60 days postpartum and declines gradually. Panel B illustrates milk production in sows that were machine milked 4 times per day. Panel C shows the yield per milking in anterior (diamonds) compared with posterior (circles) mammary glands in sows.
While there is likely some question of the accuracy of yield estimates for nondairy mammals, comparisons of peak milk yields between species give valuable insight toward physiological demands of lactation. In particular, scaling milk production relative to body weight or mammary gland weight suggests common relationships and illustrates the effect of genetic selection for milk production. Although only a small number of the known mammals have been evaluated, when data for 19 species including the mouse, hamster, rabbit, pig, human, goat, sheep, cow, and camel are used, unselected species array in a regular pattern for relationships between milk yield and body weight, energy output in milk and body weight, and mammary weight versus body weight. Equations for these relationships include the following examples (Hanwell and Peaker, 1977):

Mammary weight (kg) = 0.045 body weight (kg)0 82.
Per unit of body weight, smaller animals have higher milk yields, greater outputs of energy in their milk, and relatively larger mammary glands than other mammals. If data are averaged, the relationship between mammary gland weight and milk yield can be expressed by the following equation: milk yield (kg∕day) = 1.67 ? gland weight (kg)0,95. Because the exponent does not differ significantly from unity, on average, Ig of mammary tissue from a lactating animal produces 1.67mL of milk per day. There must certainly be some variation in this ratio, that is, variation in the degree of differentiation of the alveolar cells, contribution of accumulated secretions to weight measurements, stage of lactation, and milking or suckling patterns, but this value provides a reasonable rule of thumb for comparative purposes. It comes as no surprise to good dairy cattle managers that energy and metabolic demands on the high-producing dairy are very great. The energy and substrate demands to support increased milk production are met by a combination of increased feed intake and mobilization of body reserves. The typical dairy cow is in a negative energy balance for much of early lactation. However, when compared with smaller mammals, these demands seem relatively less severe. This is a reflection of not only demands of the suckling young but also the fact that smaller mammals typically secrete a much more energy-dense milk than cows. As an extreme case, it is estimated that the pygmy shrew (Sorex minutus), which weighs only 5g, must more than double its food consumption during lactation and eat more than four times its body weight per day to meet the additional demands of lactation. Lactation in small animals generally involves a relatively short period of intense metabolic demand, but in larger animals there is a longer period of lesser demand.
The relationship between milk yield and body weight is illustrated in Figure 18.15. The regression line for several species not selected for milk production is shown. Because certain species have been selected for high yields (dairy cows, dairy goats), values for some of these highly selected animals are also plotted as individual data points. Data points for these animals are above the calculated regression line for the nonselected species, that is, well above 95% confidence intervals, reflecting greater than expected milk production. The success of genetic selection for milk yield in dairy cows, a trait with a heritability of only 0.2-0.3, is such that a typical Holstein cow has been converted to the metabolic size of a dog and exceptional cows to the metabolic size of rodents. Concern for good husbandry practices and health of
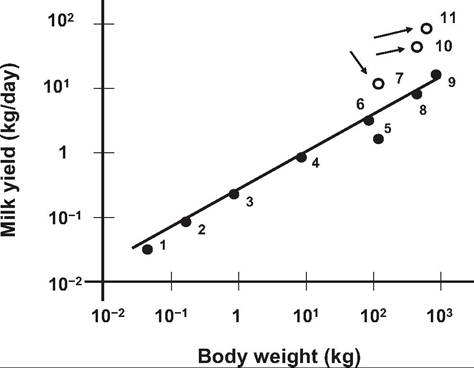
Fig. 18.15. Relationship between bodyweight and milk yield. Comparisons between dairy (open circles) and nondairy animals is illustrated. 1 - mouse, 2 - hamster, 3 = rat, 4 - fox, 5 - human, 6 = goat, 7 - dairy goat, 8 = sheep, 9 = beef cow, 10 - Holstein cow, 11 =Jersey cow. Adapted and redrawn from Hanwell and Peaker (1977).
dairy animals particularly in face of recent techniques (e.g., use of bovine somatotropin) to increase production and continuing genetic progress is justified, but it is equally important view these physiological demands on the dairy cow within an appropriate frame of reference (Mepham, 1983).
Any discussion of milk synthesis has to consider not only the nutrients necessary for milk synthesis but also the physiological adjustments necessary to supply these nutrients to the mammary gland. Requirements for high levels of milk production are staggering at first glance. For example, in the dairy cow the energy requirements for milk production can easily approach 80% of net energy of intake. Demands for lactose production can require 85% of available glucose. It has been calculated that high-producing cows must mobilize adipose tissue and body nutrient reserves equal to approximately one-third of the milk produced during the first month or more of lactation. Clearly, finely tuned coordinated interactions between all the major physiological systems are necessary for success.
Galactopoiesis and bST
Administration of bST to lactating dairy cows increases the yield and efficiency of milk production. In response to injection of bST, milk secretion increases within a day and is maximized within a week. The increased milk yield is maintained as long as treatment is continued but quickly returns to control levels when bST is discontinued. The milk yield response is dose dependent and the response curve hyperbolic. At approximately 40 mg of bST / day, nearly maximal response is obtained. Milk yield achieved with near maximal doses of bST is impressive, with increases reported as high as 30-40%. Typically bST increases milk production by 4-6 kg/day, approximately a 10-15% increase in yield. The magnitude of response to a particular dose of bST depends upon biological variation, stage of lactation, and management parameters. The bST formulation currently approved for use in the United States is a prolonged-release N-methionyl- bST (Posilac, Monsanto Co.) that was approved by the United States Food and Drug Administration in November 1993. It is administered at a dose of 500 mg per cow every 2 weeks. Package instructions are that treatment should be initiated after peak lactation at >60 days postcalving, when cows are at or near positive energy balance.
The first year after approval average milk yield increased 3 kg, and this level of increase for bST- adopting herds has been maintained. Since not all cows in the bST herds are treated, that is, some cows are not eligible < the 60 days postpartum, this is a conservative estimate of the effect of bST treatment on milk production (Bauman, 1999).
Given the voluminous literature and now very wide spread commercial use of bST in dairy cows in the United States and other countries, testing and evaluation of this technology has likely been the most intensely scrutinized new animal technology in history. Public debate about bST use was also extensive. Special interest groups and very public individuals predicted dire consequences from adoption of bST for use in dairy cows. Among concerns that had little if any scientific basis, bST approval would cause a massive reduction in milk consumption, milk price would decline, and farmers would go bankrupt. Others predicted dire animal consequences. Media coverage was extensive near the time of approval by the Center of Veterinary Medicine of the Food and Drug Administration in 1994, with more than 800 reports in the first quarter of the year. At the current time, the regulatory agencies of more than 50 countries have reviewed safety concerns and approved bST for use. It estimated that more than 3 million cows currently receive bST supplementation. In the United States, this includes animals in herds of all sizes, located in every region of the country. Scientific and anecdotal information support the view that bST is a safe, effective, and profitable management tool for the dairy farmer. Although U.S. food laws do not mandate labeling of milk from bST-treated cows because the composition of the milk from the bST cows is equivalent to that of controls, a relatively small niche market has developed for milk obtained from herds not using bST. However, since there is no chemical test to distinguish the milk from bST- and non-bST-treated cows, validation of milk from non-bST treated cows simply depends on the fact that these producers sign a certification declaring that bST is not used on their farms. This is analogous to niche markets for other organically farmed products. A recent estimate is that milk from these farms constitutes less that 1% of fluid milk sales in the United States. This suggests that the vast majority of consumers are interested in wholesome food products obtained at competitive prices. In fact during the first year after approval, milk fluid consumption in the United States increased about 1%.
Reasons for some of the rancorous debate on the approval of bST for use in dairy cows were varied but logic suggests several elements were important. This was the first proposition for widespread treatment of animals with a recombinant DNA-derived product that impacted not animal health but animal production. Second, the wholesomeness of milk and milk products seem to occupy a special position when compared with many other food materials. Third, the now roundly refuted dire predictions of consequences on animal health (Collier et al., 2001) coincided with a growing affluence and associated "greening" of attitudes among many segments of the population. Finally, popular concerns that the adoption of the technology might hasten the disappearance of family farms and change the sociology of rural farm areas were seemingly carried along on a wave of nostalgia at the time. Certainly it was an interesting process to observe. Regardless, responses of milk cooperatives, grocery chains, and other marketing concerns make it clear that decisions related to use of technologies such as bST do not necessarily follow the scientific evidence.
Mammary involution
Mammary involution can be stimulated at any stage of lactation by removal of suckling young or in dairy animals by suspending milking. Most of the detailed cellular and molecular research on mammary involution has focused on effects of acute induction of involution. However, after the peak of lactation, a gradual involution occurs, as the young are progressively weaned. In dairy cows, there is also a gradual decline in milk production with time even with regular milking. This decline is greater for cows that become pregnant soon after calving.
Cellular details of mammary involution have been most studied in rodents. Regardless of the differences in timing of events between species, that is, days to hours in rodents versus days to weeks in ruminants, involution involves apoptosis (or programmed cell death) of the alveolar epithelial cells and tissue remodeling. Apoptosis of mammary epithelial cells occurs in several phases of mammary development, not just mammary involution. These include canalization of major mammary ducts in fetal development, rapid elongation of mammary ducts via the terminal end bud, and the waxing and waning of ductular growth during estrus or menstrual cycles. Apoptotic destruction of abnormal cells at early phases of carcinogenesis is also a likely mechanism for protection against breast cancer.
In rodents, removal of the suckling young and accumulation of milk at midlactation rapidly initiates involution of the mammary gland. Marked changes in gene expression are evident within 24 hours and evidence of widespread apoptosis observed by 48 hours. For example, in rats, expression of mRNA for the caseins is reduced 95% and that of acetyl-CoA carboxylase, a key Iipogenic enzyme, 98% within 24 hours. Translational activity is also reduced. A second stage of involution is initiated between 72 and 96 hours with the activation of a series of tissue proteinases, including stromelysin 1 and 2, gelatinase A, and plasminogen activator. By this time, remodeling of the mammary gland to prepare for a new reproductive cycle is well underway. Given the rapidity and magnitude of these tissue changes, it is not surprising that involution in the mouse is only partially reversible if suckling young are returned within 48 hours. Although much of the involution process is initiated by milk stasis, the continued secretion of lactogenic hormones associated with milking or suckling can delay the process. This is shown by experiments in which suckling is allowed in only selected glands. For example, in lactating sheep, when both glands were nonsuckled for the first 15 days postpartum, mammary prolactin receptor was reduced 84%, but in the nonsuckled gland of ewes with the opposite gland having continuing suckling, stimulation the prolactin receptor was reduced only 36%. Parenchymal DNA concentration was not affected by suckling treatments; however, RNA concentration followed a pattern similar to prolactin receptor, lowest when neither gland was suckled (2.1 mg∕g), highest in suckled glands (7.4 mg∕g), and intermediate in nonsuckled glands companion to a suckled gland (3.8mg/g). Alveolar structure was maintained in all treatments but the cytological appearance of the epithelial cells reflected changes in RNA concentration. Epithelial cells from ewes with neither gland suckled were poorly maintained, with many cells engorged with secretory vesicles and lipid droplets (Box 18.2).
Involution in the ruminant mammary gland is decidedly slower with less loss of alveolar structure. After 3 days of nonmilking casein and Oc-Iactalbumin mRNA was reduced but β-lactoglobulin mRNA was unchanged. After a week, mRNA for the two milk proteins α-slcasein and Oc-Iactalbumin was dramatically lower (85 and 99%, respectively). After 4 days of nonsuckling, alveolar structure is degenerated in the rodent and apoptosis, based on the degree of DNA laddering, is near maximal. However, even in nonpregnant, lactating beef cows, alveolar structure was largely intact in the absence of suckling, with only isolated areas of tissue degeneration evident even after several weeks. Nonetheless, based on quantitative histology, well-differentiated cells, common in suckled glands, were rare in nonsuckled glands. Interestingly, regression of the gland in the absence of sucking was not uniform. Structure of parenchymal tissue distant from the teat was better maintained. Even 42 days after cessation of suckling, areas of alveolar structure were present. Overall, alveoli from lactating glands had more cells per cross section (30.4 ± 0.9) compared with 21.4 ± 0.8 in glands not suckled for 42 days. In total, these data illustrate that even a prolonged period of nonsuckling (42 days) does not result in complete destruction of the mammary alveoli. However, since total gland DNA was reduced (50-64% after 21 and 42 days of nonsuckling, respectively), cells are lost with prolonged
Box 18.2 Milk composition and milk removal Does the composition of milk influence milking or suckling capacity? As illustrated in an earlier chapter (Fig. 2.10), secretory processes in epithelial cells can be influenced by specific products made by the cells.
The disaccharide lactose appears in all milks, with the exception of those of some marine mammals. Its synthesis depends on the enzyme lactose synthetase, which is active as a complex of βl-4- galactosyl transferase and the hormonally regulated milk protein Oc-Iactalbumin. It is known that Oc-Iactalbumin evolved from lysozyme prior to the evolution of sauropsids (leading to birds and reptiles). Thus, the ability to produce lactose appeared before it was utilized as a milk component. It is likely then that many of these early milk-like secretions contained antimicrobial oligosaccharides but no lactose. Interestingly, the cellular biosynthesis of lactose in the modern mammary gland also acts to promote hydration of suckling young because of the capacity of lactose contained within secretory vesicles to attract water via osmosis, thus effectively diluting mammary secretions. It is also likely that the presence of lactose improves milk letdown and the supply of milk to the suckling young because of the production of less viscous secretions. Indeed, experiments in transgenic mice have demonstrated that when the Oc-Iactalbumin gene is silenced, the pups fail to thrive because of a failure to productively nurse (Stinnakre et al., 1994).
This suggests that there must be differences in oxytocin release, response times, or mammary architecture that allow for sufficient and efficient milk removal in mammals with milk containing as much very high fat (as much as 50%) but no lactose (fur seals and sea lions) or very low (normally occur (Box 18.3).
Because of the typical breeding pattern, little attention has been given to determining an optimal dry period for goats or sheep. However, when lactating
Box 18.3 Lactation strategies
Fur seals and sea lion (Otariidaes) have developed a novel approach to lactation and optimizing offspring survival. Mothers typically forage at sea then return with others to isolated areas to give birth and stay with their pups for a relatively short initial suckling period (~1 week); thereafter, the mothers regularly return to the sea to forage often for several days or weeks at a time. They then return to periodically suckle their pups (usually 1-3 days). In contrast with other mammals they have evolved an ability to minimize milk synthesis during forage cycles at sea so that mammary regression and involution does not occur. The milk for pups is every high in fat, ~35%. The transfer of this energy allows the pups to rapidly gain weight and lay subcutaneous fat, which insulates the pups and provides nutrition for periods when mothers are foraging. These repeated cycles of suckling and foraging continue throughout lactation (4-12 months) depending on the subspecies.
The Phocids or true seals have adopted a fasting strategy regarding lactation. Especially for the larger animals, mothers give birth and use mobilization of body reserves to supply milk for all of the lactation period. In these animals, lactation lasts for little as 4 days (4-42 days), then pups are weaned. During this time the pups grow rapidly because of the high levels of fat.
The Odobenids, the walrus, for example, use an aquatic nursing strategy. Mothers build up blubber stores, then, during a short fasting period, they give birth (typically on an ice flow), the pup suckles and bonds, and soon after it accompanies the mother into the sea to forage. Lactation can continue for 2-3 years, but by about 5 months, pups are foraging and nursing. Walrus milk is lower in fat compared with other groups so pups grow relatively slower and the physiological investment in lactation is extended.
These examples give a taste of some of the variety in lactation strategies that have evolved in various groups of mammals (Sharp et al., 2006).
goats were hormonally induced to ovulate and mated during the usual seasonal anestrus period, they entered the next lactation without a dry period and produced 12% less milk than controls. However, the data were confounded with possible effects of season. Consequently, in a second study, these researchers used a within-animal experimental design to determine the effect of drying off on lactation in goats. One gland was milked continuously, and the other dried off 24 weeks before parturition. There was no difference in subsequent milk production between udder halves. In contrast with the first study, this suggests a dry period is of no benefit. However, the dry period was relatively very long (i.e., three times the optimal length for the cow). Finally, udder-half experimental design may be less than ideal for this evaluation. Interactions of glands of differing lactational state within the udder on cell turnover and lactogenesis have not been investigated. Since prepartum milking advances lactogenesis and milk production, milking one gland may conceivably advance lactogenesis and milk production in other glands. In nonpregnant ruminants, milking delays mammary involution in other nonsuckled glands. Thus, it is possible that milking one gland during the prepartum period inhibits the ability of the opposite gland to produce maximally during the subsequent lactation, or that milk production was increased in glands milked continuously when the opposite gland was dried off. The importance of a dry period to maximize milk production in dairy goats and sheep is unsettled. Studies to determine the effects of pregnancy status, length of the dry period, and other implications of the udder-half experimental design are warranted (Wilde et al., 1999).