Overview of mammary development
Although the evolutionary origins are unclear, the mammary epithelium arises from the germinal ectoderm and the primitive mammary buds. The first indication is a slight thickening of the ventrolateral ectoderm in the embryo at about the time that limb buds begin to lengthen.
This thickened tissue is variously referred to as the mammary band, streak, or line. The cells within the mammary line condense or coalesce to form mammary buds. Each mammary bud gives rise to each of the individual mammary glands. In ruminates, the mammary buds are closely grouped so that the developing mammary glands are oriented in an udder, for example, two glands in goats and sheep but four in cattle. In the bovine, the mammary buds appear at about day 40 of gestation. By day 80, the teat and primary sprout form. The primary sprout gives rise to the teat cistern. Secondary sprouts occur by day 90, and by day 100 the primary and secondary spouts are canalized. At birth, the teat, teat cistern, and gland cistern are formed. In most species, mammary structure at birth is similarly rudimentary.Figure 18.1 illustrates the epithelial portion of the mammary gland of a mouse at 1 week of age. Mice of
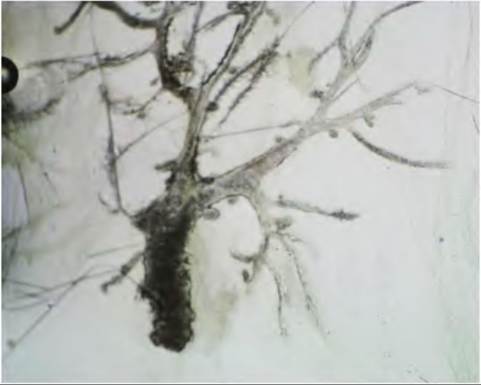
Fig. 18.1. Rudimentary mammary gland of the 1-week-old mouse. The nipple and primary mammary sprout are oriented at the lower portion of the figure. Other secondary sprouts radiate as if they were bare tree branches into the surrounding fat pad.
course do not have a teat or gland cistern, but the structure of the nipple and branches from a number of secondary sprouts are evident. For mammals without the mammary glands arranged into an udder, a teat or gland cistern is absent, but there is a nipple and a cluster of primary and secondary sprouts for each gland.
The mammary gland is one of only a few tissues in mammals that repeatedly undergo growth, functional differentiation, and regression.
This is one of the reasons for very great interest in study of the mammary gland. The term mammogenesis refers to the development of mammary gland parenchymal structures. In usual circumstances, studies of mammogenesis are focused on the very large changes in the mammary gland, which begin around the time of puberty or more likely during pregnancy. However, the foundation for the dramatic mammogenesis during these stages begins when the animal is an early fetus. Fundamentally, mammary development can be developmentally considered a joining of the epithelial ectoderm and the underlying mesoderm.Across species, there are dramatic differences in milk composition. Milk from Holstein cows (the source of the majority of milk for human consumption in Western societies) has about 3.2% protein, 3.4% fat, and 4.6% lactose. In contrast, hooded seals produce milk with about 6.0% protein, 50% fat, and virtually no carbohydrate. Much of this variation reflects evolution- induced responses, which provide the best stratagem for offspring survival. Because the seal pups are born on potentially unstable pack ice, they must rapidly gain sufficient strength and insulation to survive. In fact, mothers suckle their pups for only 4 days during this period, but pups can double their 20-kg birth weight. While the hooded seal has the shortest lactation of any known mammal, the high-fat milk provides the pup with the energy and metabolic water necessary for an abrupt introduction into a polar environment.
Prepubertal and postpubertal mammary development
Generally, there is little or no true Iobuloalveolar development before conception. This period is associated with creation of a framework to allow proliferation of the secretory alveolar cells needed for lactation. The period is generally considered a time during which the duct system is extended and growth of the adipose and connective tissue increases. Ductular growth is limited for dogs, cats, and rabbits but is more extensive for rats, mice, the rhesus monkey, cattle, and sheep.
Since it is expected that most organs in growing animals would also grow in concert with rate of overall body growth, the regulated, cyclic development of the mammary gland can be difficult to study under these conditions. One approach is to evaluate mammary development in terms of relative growth and to ask if growth of the mammary gland fits the law of simple allometry, y = bx“. A usual approach is to log transform variables associated with body and tissue growth and use linear regression analysis to calculate the equilibrium constant (α), which relates the difference in growth rate of the organ under study to the growth of the body as a whole. Simple body weight or body weight273 (to approximate surface area) is usually used as the independent variable in these analyses. The dependent variable (mammary gland area, mammary gland weight, DNA content, weight of parenchymal mass) serves as an index of the growth of the mammary gland. When α = 1, the growth is said to be isometric. If a > 1, the growth rate is positively allometric (simple allometry), but an α < 1 indicates growth rate of the organ is negatively allometric (enantiometry).
Data for rodents, cattle, and sheep demonstrate that from birth until before puberty, the mammary gland grows only somewhat faster than the rest of the body. Thereafter, the rate of mammary gland growth becomes markedly allometric usually through the first few estrus cycles. Growth then reverts to a period of isometric growth that also waxes and wanes during the course of the estrus cycle. As demonstrated for heifers, the classic study by Sinha and Tucker (1969) shows that positive allometric begins at about 3 months of age and continues until about 9 months of age. Specifically, when mammary development was expressed as log of mammary DNA and body growth as log of body weight, the equilibrium constant (α) averaged 1.6 in the period from birth to 2 months, 3.5 from 3 to 9 months, and 1.5 between 10 and 12 months of age (Fig.
18.2). These data indicate thatthe mammary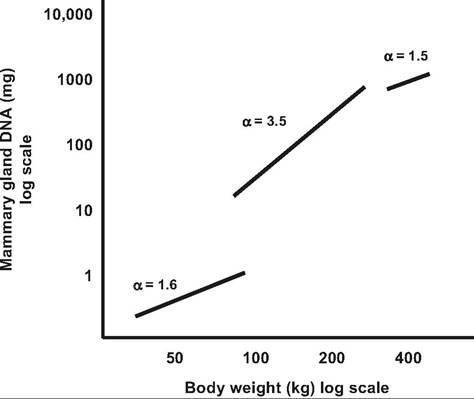
Fig. 18.2. Relative mammary growth in heifers. The data illustrate the relationship between mammary DNA and body weight in Holstein heifers. Segments illustrate periods from birth to 2, 3, to 9, and 10 to 12 months of age. Reproduced from Sinha and Tucker, 1969.
gland grows somewhat faster than the body during the early postpartum period but dramatically faster between 3 and 9 months of age.
Although the absolute amount of mammary growth during the period prior to conception is only a fraction of the mature mammary gland late in gestation, there is compelling evidence that alterations in mammary growth during the peripubertal period can affect subsequent mammary function. For example, rapid prepubertal weight gain inhibits subsequent mammary parenchyma tissue growth and can reduce subsequent milk production (Sejrsen and Purup, 1997; Sejrsen et al., 2000). Not surprisingly, very few cattle studies have considered changes in mammary development in the period shortly after parturition. While minor in terms of absolute mass, our observations of mammary glands of Holstein calves between 1 and 3 months of age (Capuco and Akers, 2010) suggest that parenchymal tissue per gland increases approximately 60-fold in only a few weeks. It is intriguing to consider that growth during even this very early postnatal period may also be critical to the success of lactation. Indeed, it may be that induction and/or regulation of the appearance of mammary stem cells will be important in new tools and techniques to regulate and control mammary development and function in the future (Capuco et al., 2012; Ellis et al., 2012).
Figure 18.3 illustrates various stages of mammary development using the mouse mammary gland as an example. Small rodents are especially useful for this purpose because an entire mammary gland can be removed, spread onto a microscope slide, fixed, defatted, and stained.
Observing structural differences in mammary glands from various physiological stages quickly illustrates the dramatic changes in the development of the mammary epithelium as the gland prepares for the onset of lactation. However, a word of caution is advised. Although the fundamental developmental processes are similar in rodents and other species, there are clearly substantial differences in the tissue composition of the mammary gland in rodents compared with other species as well as differences in the pattern of ductular development.Compared with the rodent pattern of development where the end buds allow for filling of the entire mammary fat pad during the prepubertal period (Fig. 18.3), the developmental pattern in the ruminants is more compact. Specifically in rodents, as the epithelial ducts develop, they are surrounded by stromal cells, and the stromal tissue contains a sea of adipocytes. A higher power view (Fig. 18.4B) of a similar area from the mammary gland of a prepubertal heifer illustrates several epithelial structures, which radiate from a mammary duct. Pre- and postpuberty cross-sectioned ducts often demonstrate a scalloped appearance, suggesting a complex tubular structure. Indeed, three- dimensional, computer animations prepared from
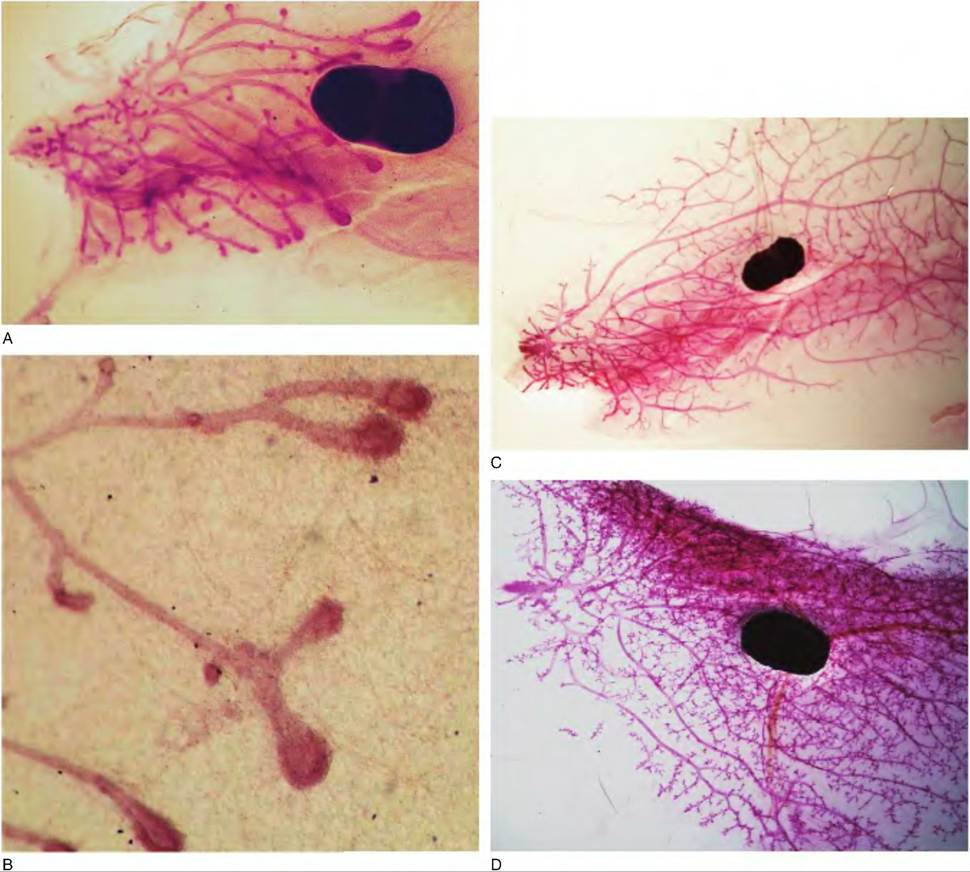
Fig. 18.3. Physiological stages and mammary development. These images illustrate a series of whole mounts prepared from an inguinal mammary gland of mice at various stages of development from the prepubertal period into lactation. Panel A shows the gland of a 4-week-olcl mouse. Ducts have elongated from the region of the primary sprout and nipple (left) to just beyond the lymph node. This progressive growth depends on the highly mitogenic end buds, which appear as enlargements at the ends of the ducts. Panel B shows enlarged end buds. C shows a gland from a mouse at about 40 days of age. The mammary fat pad is largely filled with mammary ducts but notice the end buds have now regressed.
D shows the mammary gland of a mouse after the onset of estrus cycles. The number of branches has increased and alveolar buds have begun to appear. E shows enlarged alveolar buds. F provides an image of just a portion of the mammary gland on day 12 of gestation. Alveoli are now beginning to develop and the gland becomes thickened so that images of tissue structure are only possible at the thinner margins of the gland. G provides an enlargement of an area of developing alveoli. H shows the margin of the mammary gland on day 4 of lactation. Because of the marked increase in alveolar growth and accumulation of secretions, the rounded structure of clusters of lobules and alveoli is evident at the thinner margins of the whole mount.serial sections of prepubertal bovine mammary parenchymal tissue elegantly confirm this tissue architecture (Capuco et al., 2002). This suggests that in the ruminant, in contrast with rodents, the gland is not filled with elongated ducts during the prepubertal period waiting for subsequent development of side branches. To use a plant analogy, in the peripubertal rodent, widely spaced mammary ducts fill the mammary fat pad like the bare branches of a tree. In the peripubertal ruminant, closely packed ducts radiate from the gland cistern in broccoli-like fashion, but the ducts generally fill only a fraction of the mammary fat pad.
As the gland continues to develop, the relative tissue area occupied by the epithelium increases at the expense of the surrounding stromal tissue. This is especially evident with the growth of alveoli during gestation. It is also evident that much of the tissue area in histological sections of mammary tissue taken from cows in
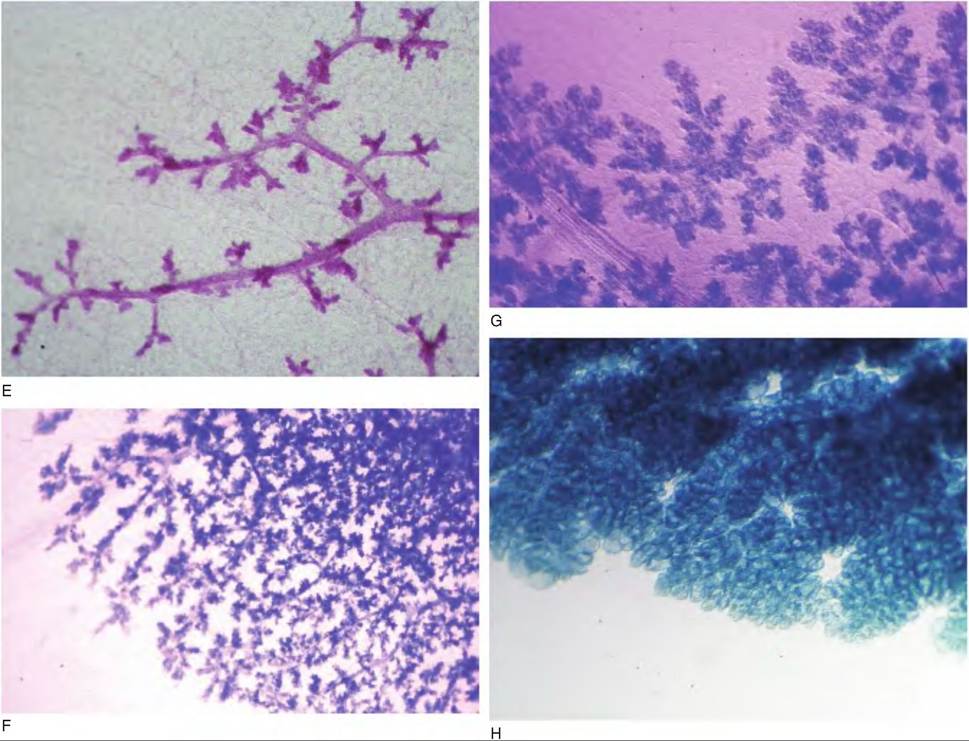
Fig. 18.3. Continued
late gestation and lactation is also occupied by Iumenal space. This does not mean that there is necessarily a loss of stroma cells as the gland develops but rather that there is a dramatic rearrangement of cells and tissue elements so that that the stromal cells are less evident. As gestation advances, clusters of alveolar structures appear as scattered, round islands, until late in gestation when histological fields are filled with closely packed alveoli. Under normal circumstances, true alveoli are not formed until conception. Variation among species reflects the number of estrus cycles that occur prior to conception. Using cattle as an example, in the early stages of pregnancy the duct system continues to develop with appearance of a rudimentary Iobuloalveolar system by about 5 months of gestation.
Mammary development during pregnancy
It is estimated that 94% of mammary development for the hamster takes place as pregnancy advances. Estimates for other species range from 78% for the mouse and sheep to 66% in rabbits and 60% for rats. By far the greatest promoter of natural mammary growth or mammogenesis is pregnancy and associated hormonal and physiological changes. With influence of pregnancy, mammary growth is reinitiated after reversion to isometric growth following puberty. This growth can be described by an exponential equation with the following form: Y = au where Y = mammary size, f = day of gestation, and the terms a and b are constants. Such equations have been developed to model mammary growth in cows, goats, and guinea pigs. The term a is mammary gland mass or size at the beginning of gestation, and the term b is the first-order rate constant. The time necessary for the mammary gland to double is equal to ln2∕b (Sheffield, 1988).
Measurement of mammary DNA or weight of dissected parenchymal tissue is useful to quantify rates of mammary growth, but these techniques do not distinguish differences among cells types. Problems of trying to distinguish between stromal and parenchymal tissue are especially difficult in rodents. For
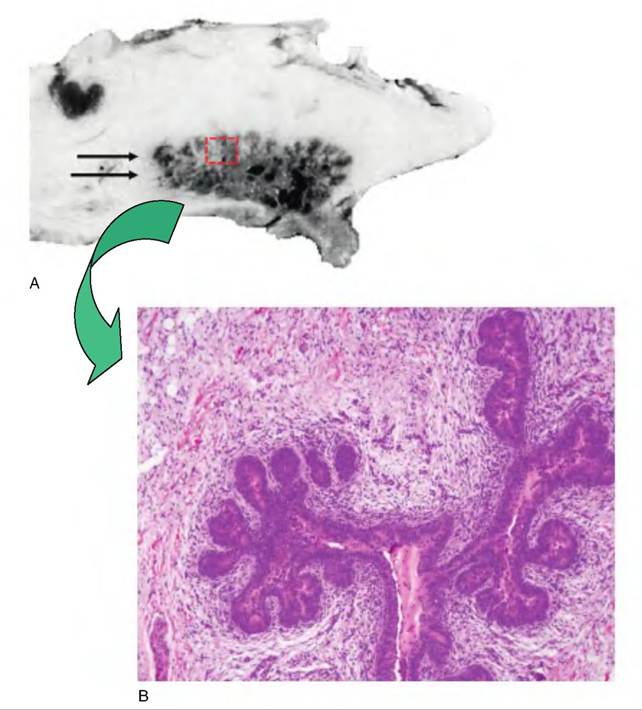
Fig. 18.4. Ruminant mammary development. Panel A illustrates a mid-saggital section through the udder of a prepubertal ewe lamb and panel B a histological section of mammary parenchymal taken from a region similar to that outlined by the dashed box and indicated by the double arrow.
Table 18.2. Effect of gestation on mammary growth, histology, and epithelial cell labeling index in ewes.
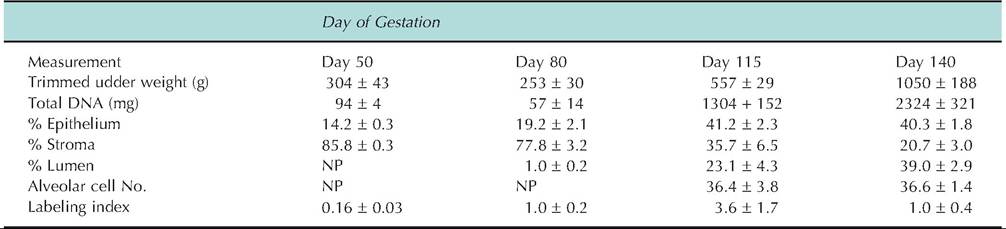
Data adapted from Smith et al. (1987). NP indicates not present; specifically alveoli appeared between day 80 and 115 of gestation. Labeling index indicates the percentage of epithelial cell nuclei that incorporated tritiated thymidine following a one-hour incubation of tissue explants.
example, both wet weight and defatted dry weight of the inguinal mammary glands of rats are reported to vary from 50% to 120% during pregnancy but total mammary DNA changes 200-300%. Since total DNA is a reflection of cell number, changes in weight alone underestimate cellular development. Table 18.2 illustrates changes in mammary development of ewes during gestation. Changes in weight, total DNA, or percent of epithelium show a marked increase in parenchymal tissue between day 80 and 115 of gestation. However, between day 115 and 140, quantitative histology (percent of epithelial area) alone suggests there is no continued parenchymal development. This is clearly not the case, since both tissue weight and total DNA double. Lack of change in epithelial area between day 115 and 140 is a reflection that alveoli are present at these stages of gestation, but an increase in lumen and decrease in stromal tissue area reflect accumulation of some secretions and compression of stromal tissue between alveoli. In addition to the appearance of alveoli during gestation, the cells undergo a progressive structural and biochemical maturation as parturition approaches. The increase in DNA labeling index on day 115 reflects the rapid growth during this period.
Mammary growth as measured by total DNA continues during the early days of lactation in rats, rabbits, guinea pigs, and mice. In the rat, for example, as much as 26% of the total mammary growth occurs during lactation. In the mouse there is a transient surge in mammary cell proliferation 2-3 days postpartum. In fact, it is reported that the cell population doubles between the last day of gestation and day five of lactation. The guinea pig is especially interesting in that there is little change in total mammary DNA during gestation but a dramatic increase within 2 days of the birth of the pups. However, if suckling is not permitted in these species, the increased growth just after parturition is prevented. This suggests that signals associated with suckling or milking are important for growth especially in early lactation. Regardless, it seems clear that once lactation is established, the rate of mammary cell proliferation is markedly lower than during other stages of mammary development. However, recent reports show that several weeks of "extra" milking stimulation within the first 2 months of lactation in dairy cows increase subsequent milk production when milking frequency is returned to normal. Mechanisms for the effect are unknown, but the fact that it continues after treatment supports the idea that it may involve recruitment of additional well-differentiated cells. This might reflect overt proliferation of additional secretory cells or perhaps enhanced functional differentiation in a population of preexisting nonsecretory cells (Bar-Peled et al., 1995). This effect contrasts with the galactopoietic treatment of cows with bovine somatotropin (bST) in that milk production is dramatically increased during the period of treatment but returns to control levels when treatment is discontinued. This pattern of response has been interpreted to indicate that bST alters metabolism rather than cell proliferation. This is clearly an area of continuing research interest.
In dairy species, it is usually concluded that there is normally little mammary growth during established lactation. However, comprehensive data for early lactation is lacking. The concentration of DNA in mammary parenchyma is relatively unchanged in early lactation in sheep, goats, or cows, but it is risky to interpret this as a lack of mammary growth since it does not evaluate the entire mammary gland. Total parenchymal DNA doubles between 2 weeks before and after parturition (27.9 vs. 46.0 g), but these data do not determine if the growth occurs before or after parturition. Rates of thymidine incorporation are very low for mammary tissue taken from lactating ruminants, but mitotic cells are sometimes observed (Capuco and Akers, 1999).
In lactating goats, unilateral inhibition of secretion in one gland of goats results in a compensatory increase in milk production of the other gland. There are also reports of increased milk production in cows by uninfected quarters among cows with mastitis. The relative contributions of hypertrophy and hyperplasia for this effect are unknown. However, in lactating beef cows, covering one-half of the udder to prevent sucking increased thymidine incorporation in lactating mammary glands compared with control glands of cows that continued suckling in all glands. Morphologically, lactating tissue from control and compensatory treatment cows was indistinguishable and the tissue did not appear consistently different between zones within lactating quarters. About 40% of the parenchymal tissue consisted of closely packed alveoli. In some areas, the secretory cells appeared highly differentiated similar to cells from dairy cows. Remaining parenchymal tissue contained alveoli more widely scattered in the stromal matrix, and the cells were less well differentiated. Labeled cells were observed in both regions but appeared more frequently in the less well-differentiated tissue (Akers et al., 1990).
Figure 18.5 shows changes in the growth of mammary parenchymal of beef and dairy heifers during pregnancy and into lactation. Comparisons between panels A and B illustrate those measures of udder parenchymal mass alone but can be misleading. Specifically, based on udder parenchymal mass, there is marked growth of parenchymal tissue between day 260 and day 49 of lactation in both Holsteins and Herefords (Fig. 18.5A). In contrast, when udder growth is evaluated based on parenchymal DNA (Fig. 18.5B), the marked differences between breeds remain, but little change occurs between day 260 of gestation and day 49 of lactation in Holsteins. This is likely explained by the increased accumulation of secretions in the Holstein heifers compared with the Hereford heifers. When lactation performance is considered, it is clear that differences in milk production (3.5 vs. 20.3 kg∕ day) are explained by changes in udder parenchymal tissue mass and function. For example, total RNA as well as RNA/DNA ratio is greater in Holstein than Hereford heifers. Interestingly, ability of mammary explants from lactating animals to secrete Oc-Iactalbumin (a specific milk protein) closely mirrored (57 vs. 289 ng per mg of tissue per 24 hours) the corresponding 5.8- fold difference in daily milk production. The relative failure of cytological differentiation (Akers et al., 2006) in mammary tissue of beef compared with dairy heifers is puzzling since milking stimulation and manage-
merit was identical between breeds in this study. Is it possible that selection for increased milk production (in dairy cattle) has allowed for the maximization of differentiation signals during the critical periparturient period?
Realization in the early 1960s that the DNA content of cells is essentially constant (with the exception of
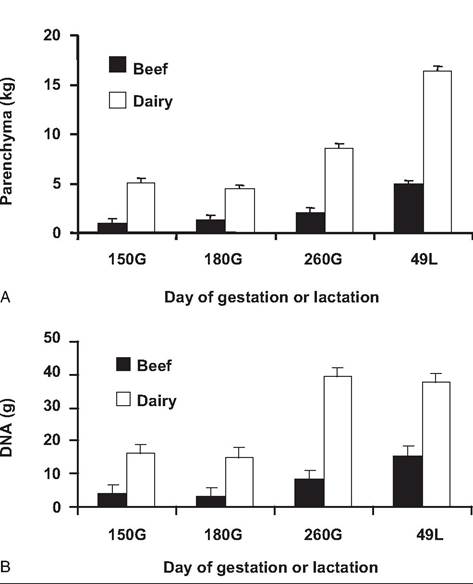
Fig. 18.5. Mammary growth during gestation in cattle. Chart A illustrates the growth of the mammary tissue of beef and dairy heifers during pregnancy and lactation based on weights of parenchymal tissue dissected from udder slices obtained at slaughter. Chart B provides similar data, except growth is reflected by changes in total parenchymal tissue DNA. Both measures demonstrate a consistent advantage for the Holstein compared with Hereford heifers. However, the increased weight of parenchymal mass between day 260 prepartum and day 49 of lactation for Holsteins is not reflected in DNA. This suggests that much of the increase in Holsteins reflects onset of milk secretion. Figure adapted from Keys et al. (1989).
the generally small proportion of cells that are undergoing DNA synthesis in preparation for cell division at a given moment) ushered in a host of studies to estimate mammary cell number based on total DNA content. This method is especially valuable when combined with careful dissection of the mammary gland to distinguish the parenchymal portion from the stromal tissue of the mammary gland. Even with careful dissection of the mammary gland to remove apparent connective tissue, there are clearly nonglan- dular cellular elements, that is, blood vessels, lymphatic vessels, nerves, fibroblasts, adipocytes, and white blood cells which contribute to the DNA content of the parenchymal tissue compartment. Regardless, classic studies in a variety of lactating species give direct evidence that the number of mammary epithelial cells is proportional to milk production. The correlation between total parenchymal DNA and milk production averages about 0.85. Consequently, any activity that reduces the number or function of the mammary alveoli, will also reduce milk production (Tucker, 1981,1987).
In addition to the nonsecreting epithelial cells and various stromal cells, extracellular secretions, that is, proteins and proteoglycans that surround the mammary ducts and alveoli, are also critical for development. Collagen is the major extracellular protein component of the mammary stromal tissue. Moreover, the amino acid hydroxyproline is a specific and major component of collagen. Thus, assay of tissue content of hydroxyproline provides a quantitative measure of stromal tissue. When coupled with measurement of fat, the relative amounts of connective tissue associated with the parenchymal tissue can be estimated.
Data in Table 18.1 and Table 18.3 illustrate the dramatic changes in mammary growth from birth to lactation in Holstein heifers and crossbred ewes. Measured as trimmed udder weight or parenchymal DNA, mammary growth is greatest during gestation. However, relative lack of change in DNA from late gestation into lactation compared with trimmed udder weight suggests that DNA is a better measure of cell growth, since increased weight may be accumulated secretions.
Table 18.3. Mammary parenchymal growth in heifers and ewes.
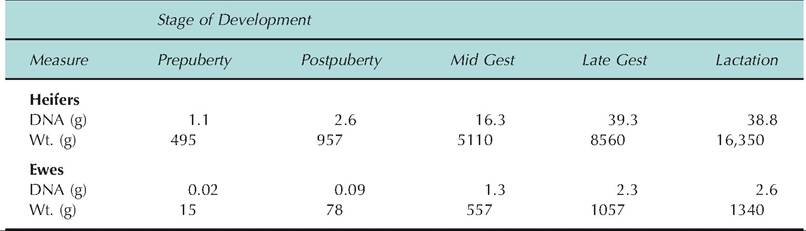
Data adapted from Sejrsen et al. (1982, 1986), Smith et al. (1989), Keys et al. (1989), and McFadden et al. (1990).
Measurement of parenchymal tissue RNA and/or protein indicates synthetic capacity and is useful to evaluate the fully developed mammary gland. The reason for this is simple. As the secretory cells differentiate in concert with parturition, there is a marked increase in the presence of rough endoplasmic reticulum and corresponding production of mRNA for specific milk proteins. Consequently, onset of milk synthesis and secretion corresponds with a marked increase in the mammary tissue RNA. On a per-cell basis, increased synthetic capacity can be evaluated by comparing the ratio of RNA/DNA in mammary parenchymal tissue. Late in gestation, after alveoli have formed but before the onset of copious milk synthesis and secretion, this ratio is generally about one. During established lactation, the RNA content of the secretory cells increases dramatically so that this ratio more than doubles. Changes in parenchymal tissue protein follow a similar pattern; however, care must be taken to account for the milk protein, which may be trapped in the tissue. Finally, any of these measures are more useful if they can be applied on a whole mammary gland basis. For rodents or other animals, in which an entire mammary gland (or parenchymal tissue) can be isolated and sampled, these measures can be related to milk production.
Alveolar cell differentiation and lactogenesis
A common biological theme is that structure follows function. The structural differentiation of the alveolar cells around the time of parturition illustrates this principle. As parturition approaches, these cells become polarized, with appearance of abundant arrays of RER, numerous mitochondria, and competent tight junctions just as full-scale milk synthesis and secretion is initiated. This process is called lactogenesis. Two events are critical. The structural differentiation of the alveolar epithelial cells allows for packaging of milk components for secretion while biochemical differentiation allows for synthesis of milk constituents. We will begin by reviewing some of the structural changes.
The mammary gland begins to reach its mature structural state late in gestation; that is, alveoli and mammary ducts are in place. Indeed, soon after the alveoli are formed, some accumulation of secretions begins. For example, in cows during first gestation, small but variable volumes of secretions can be expressed from the mammary gland several months prior to parturition. Prepartum milking of dairy heifers is sometimes initiated during the month before parturition as a management technique to relieve intramammary pressure. Irrespective of prepartum milking, secretions obtained prior to calving are generally high in protein and low in lactose compared with normal milk but with relatively small concentrations of specific milk proteins. Because extensive removal of mammary secretions prior to calving can alter the course of mammary development, that is, premature mammary cell differentiation, it is recommended that prepartum milking once initiated should continue until onset of normal milking at calving.
High concentrations of protein in secretions obtained prepartum in cows reflect the accumulation of immunoglobulins transferred into secretions from the bloodstream. As the alveolar cells differentiate, accumulation of immunoglobulins is reduced. However, the accumulated immunoglobulins in the secretions obtained with the first milking or suckling postpartum, that is, colostrum, provide passive immunity to the offspring. This is critical for those species that lack immunoglobulin transfer to the fetus in utero. With the onset of regular milking or suckling, the composition of mammary secretions progressively changes to reflect normal milk composition.
Figure 18.6 illustrates the major structure of a mammary alveolus, while Figure 18.7 and Figure 18.8
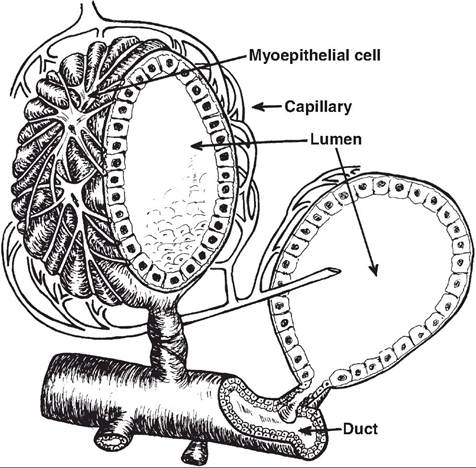
Fig. 18.6. Mammary alveolus. This diagram illustrates the three dimensional structure of the mammary alveolus. The hollow center of the alveolus provides a space for the accumulation of milk components that have been synthesized and secreted by the secretory cells that compose the internal wall of the structure. The outside of the alveolus has a network of myoepithelial cells that contract in response to release of oxytocin at the time of milking. This forces stored milk into the terminal duct, which exits the lumen of the alveolus. The milk progresses through larger ducts to be emptied at the nipple or teat end. This figure was reprinted from Larson (1985).
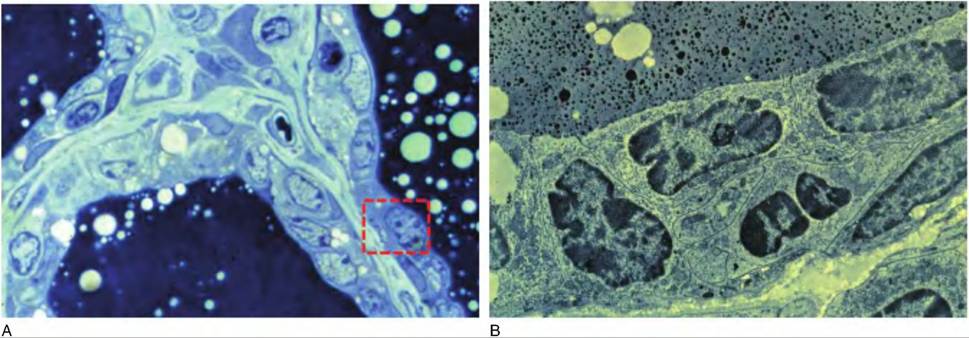
Fig. 18.7. Prepartum alveolar structure. Panel A illustrates a portion of three different alveoli from the mammary gland of a cow about 2 weeks prior to calving. This light microscope image demonstrates the relative lack of differentiation of the epithelial cells. However, the luminal spaces are darkly stained because of accumulation primarily of colostrum. Panel B illustrates the ultrastructure of cells similar to those outlined the dashed box in panel A. Notice that most of the cell area is occupied by the cell nucleus. Structurally these cells are poorly differentiated.
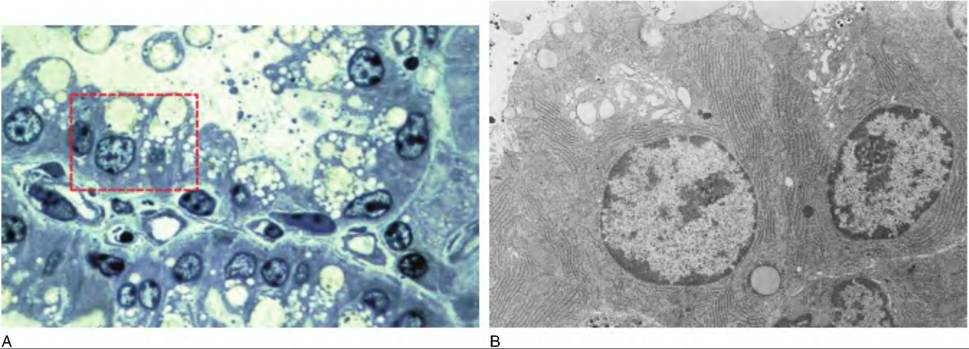
Fig. 18.8. Postpartum Alveolar Structures. Panel A illustrates a portion of two different alveoli from the mammary gland of a cow about two week after calving. This light microscope image demonstrates very well differentiated epithelial cells. The nuclei are generally rounded and basally displaced in the cell. There is also abundant RER indicated by the staining of the basolateral area near the nucleus. The apical ends of the cells have a Iacy appearance because of the abundance of Golgi, secretory vesicles, and lipid droplets. Panel B illustrates the ultrastructure of cells similar to those outlined the dashed box in panel A. Notice the rounded nuclei, numerous arrays of RER and Golgi.
illustrate the dramatic change in alveolar cell structure that accompanies lactogenesis. Much of this change is coordinated by alterations in circulating hormones and changes in hormone signaling on mammary target cells.
Ultrastructure of the mammary epithelium during lactation has now been documented in many species. The percentages of various cellular organelles within the cytoplasm of alveolar secretory cells from lactating rats and cows are illustrated in Table 18.4. This generally uniform structure includes basal and paranuclear cytoplasm occupied by parallel arrays of rough endoplasmic reticulum (RER). The supranuclear Golgi apparatus typically consists of stacks of smooth membranes whose terminal cisternae release casein- and Table 18.4. Ultrastructural analysis Ofcytoplasmic organelles in well-differentiated mammary epithelia from lactating rats and dairy cows.
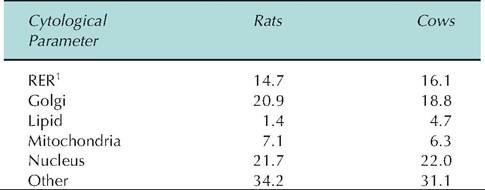
[1]Rough endoplasmic reticulum.
Data are expressed as mean percentage cytoplasm occupied by each cytoplasmic component. Data are adapted from Nickerson and Akers (1984).
Iactose-Containing secretory vesicles. Lipid droplets and secretory vesicles seemingly fill the apical ends of the cells, and microtubules are most frequently observed oriented perpendicularly with respect to the apical plasma membrane. Mitochondria and free ribosomes are abundant throughout the basal-lateral cytoplasm. The basal plasma membrane is often thrown into complex folds, believed to indicate active pinocy- tosis. Myoepithelial cells frequently occur interspersed between the basal plasma membrane and the basal lamina that forms a loose barrier between the simple cuboidal epithelial cells and the underlying stromal tissues. It is now apparent that parenchymal tissue from glands of high-yielding animals have an abundance of structurally highly differentiated polarized alveolar cells.
Hormonal control of lactogenesis and lactation
The endocrine system perhaps more than any other physiological system plays a central role in all aspects of mammary development (mammogenesis), onset of lactation (lactogenesis), and maintenance of milk secretion (galactopoiesis). Lactogenesis is frequently described as a two-stage process. Stage I consists of limited structural and functional differentiation of the secretory epithelium during the last third of pregnancy. Stage II involves completion of cellular differentiation during the immediate periparturient period coinciding with onset of copious milk synthesis and secretion. During lactation, the secretory cells synthesize and secrete copious amounts of carbohydrate, protein, and lipid. Production of this complex mixture of nutrients depends on coordination between biochemical pathways to supply metabolic intermediates and secretory pathways for secretion. For example, the disaccharide lactose is the predominant milk sugar. The enzyme complex necessary for lactose synthesis, membrane-bound galactosyltransferase and the whey protein Oc-Iactalbumin, combines in the Golgi apparatus to form lactose synthetase, which links glucose and galactose-producing lactose. Activation of the Oc-Iactalbumin gene and synthesis of Oc-Iactalbumin is most closely associated with stage II of lactogenesis.
Experiments beginning in the 1920s (Stricker and Grueter, 1928) showed that milk secretion could be induced in virgin rabbits by injecting a pituitary extract. In 1933, Riddle et al. purified the protein responsible for the milk secretion response observed by Stricker and Grueter; they named it prolactin (Prl). Even now, the widely touted and utilized galactopoietic effect of somatotropin to increase milk production in lactating dairy cows has its foundation in studies by Asimov and Krouze in the 1930s. They showed that injections of pituitary extracts consistently increased milk production in lactating cows. Scientists describing and quantifying the potent effects of the pregnancy on mammary growth and changes in the mammary gland at puberty spurred others to isolate and identify the steroid hormones estrogen and progesterone. Advances in purification techniques and understanding of steroid hormone chemistry allowed further studies leading to the production of these steroids for widespread animal testing.
Although the existence of mammogenic and lactogenic substances from the pituitary had long been known, the efforts of C.H. Li and colleagues in the 1940s to purify larger quantities of Prl and growth hormone (GH) were essential (Riddle et al., 1933). Soon thereafter, specific roles for these hormones in the regulation of mammogenesis in rodents were delineated in classic ablation replacement experiments (Lyons et al., 1958; Nandi, 1958). In an extensive series of studies, triply operated (adrenalectomized, ovariec- tomized, and hypophysectomized) rats and mice were treated with various combinations of purified hormones see if normal mammary development could be restored. Injections of estrogen and GH together caused proliferation of mammary ducts. However, treatment with estrogen, progesterone, Prl, and GH were needed for Iobuloalveolar development. The maximum ductular and Iobuloalveolar development, although still less that in pregnancy, was obtained in animals also given glucocorticoids. For some strains of mice, GH and Prl were both capable of stimulating Iobuloalveolar development.
British researchers, focused on efforts to improve and maintain milk supplies during World War II, initiated many endocrine studies on mammary development and function in dairy animals (Cowie et al., 1980). For example, the effects of estrogen and progesterone on mammogenesis were extensively evaluated in attempts to induce lactation in nonpregnant animals. Although difficulties with needed surgeries and expense continue to limit use of ablation replacement experiments to study mammary development and function in cattle, Cowie et al. (1966) studied Iiypophysectomized- Ovariectomized goats and showed that mammary development comparable to mid-gestation could be obtained in animals treated with a combination of estrogen, progesterone, Prl, GH, and ACTH. Such experiments served to confirm that at least general effects attributed to these hormones on mammary development in rodents applied to mammary development in dairy animals (Asimov and Krouze, 1937; Purup et al., 1993).
Although it should be clear that mammogenesis involves more than increased secretion of estrogen and progesterone, which occurs during pregnancy, this is nonetheless critical. During the estrus cycle, estrogen concentrations increase as a consequence of follicle growth, but with the appearance of the dominant follicle and ovulation, concentrations of estrogen decline and progesterone concentrations increase. Since estrogen and progesterone are both important for final duct growth and Iobuloalveolar development, the lack of a sustained simultaneous increase of both steroids during the estrus cycle likely explains the lack of marked parenchymal tissue development at this time. Furthermore, few of the studied species show evidence of Iobuloalveolar development prior to conception. With conception, the corpus Iuteum is maintained and along with increasing production by the placenta in many species, blood concentrations of progesterone are elevated throughout gestation. Concentrations of estradiol are higher (relative to the estrus cycle) with a gradual increase during gestation until a more dramatic increase during the final few weeks before birth. Consequently, concentrations of estrogen and progesterone are both simultaneously elevated during much of gestation, especially so during the later portion of gestation. This is believed to be responsible for much of the mammary growth during gestation. Prl and possible prolactin-like activity associated with secretion of placental lactogen is also important in mammogenesis in some ruminants (sheep and goats). In cattle, Prl is most likely a permissive agent for the mammogenic effects of the steroids and other growth factors. There are no specific changes in secretion of Prl in cattle associated with mammogenesis and Iobuloalveolar formation during pregnancy.
An interesting accidental discovery reported by Smith and Schanbacher (1973) created a flurry of activity on hormonal induction of lactation in cattle. These researchers were studying the effects of steroids on secretion of immunoglobulins into mammary secretions of nonlactating cows, that is, animals that had failed to conceive. They observed that injections of estradiol-17-β and progesterone for only 7 days, at doses that mimicked blood concentrations in animals near calving, caused the udders of some of the animals to "bag up." When these animals were milked, the initial colostrum-like secretions rapidly gave way to secretion of milk. As reviewed (Akers, 1985), subsequent studies showed that lactation could be induced in about 70% of these nonpregnant cows and that milk yields for the successful animals averaged 70% of normal. Others found positive correlations between the success of induced lactations and concentrations of Prl and well as improved yields when cows were also given drugs to induce Prl secretion. Also, greater milk yields measured for cows induced into lactation during the spring and summer was attributed to higher concentrations of Prl in serum compared with cows treated during winter months.
Although changes in blood concentrations of mammogenic hormones are important in explaining changes in mammary growth, changes in tissue sensitivity and availability of biologically active hormones are also important (Purup et al., 1993,1995). In circulation, the steroid hormones are bound to transport proteins. Even in tissues, the steroids can become sequestered with cellular lipids and therefore effectively unavailable (Capuco et al., 1982). This effect, coupled with a decrease in progesterone receptor concentration and a change in isoforms of the receptor, explains the disappearance of the negative effect of progesterone on lactogenesis at the time of parturition in cattle. This illustrates the concept that the biological effectiveness of circulating hormones may change independently of the total blood concentration of the hormone. In addition, changes in expression, synthesis, or availability of hormone receptors in target cells also clearly impact biological response. While there is little data for expression of estrogen or progesterone receptors in bovine mammary tissue during mammogenesis, data from assay of ovine mammary tissue shows that expression of the progesterone receptor occurs in close correspondence with appearance of Iobuloalveolar development (Table 18.2). Serum concentrations of progesterone are consistently elevated during gestation with higher concentrations in later stages of gestation and in ewes with more than one fetus. Table 18.5 illustrates changes in serum concentrations of some important mammogenic hormones during gestation as well as mammary tissue receptor concentration.
Table 18.5. Effect of stage of gestation on serum concentrations of mammogenic hormones mammary tissue receptor concentration.
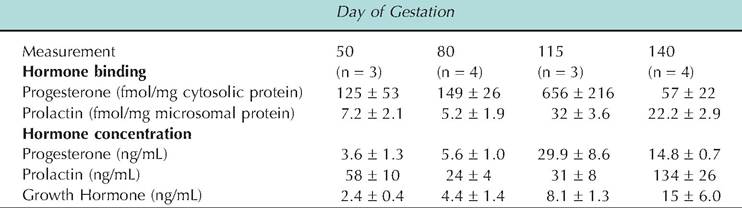
Data adapted from Smith et al. (1987, 1989).
As its name might suggests, Prl has undoubtedly been the most intensely studied hormone related to lactation and mammary growth. Despite understanding that the presence of the pituitary is essential for normal mammary development, whether Prl or GH is predominant in mammogenesis is not clear. The answer to this question is likely species dependent.
In addition to the ovary and corpus luteum, the placenta produces estrogen, progesterone, and a prolactin-like hormone called placental lactogen (PL). PL was first recognized by its Prl-Iike biological effects. However, the PLs of many species have both GH and Prl-Iike activities. For example, Prl and nonprimate GH molecules interact only with their specific receptors, called lactogenic and somatogenic, respectively. Human GH, in contrast, recognizes both receptor classes. This coincidently probably explains the inappropriate breast development sometimes observed in human males with pituitary tumors that over produce GH. Both ovine and bovine PL behave like human GH; they compete for both somatogenic and lactogenic receptors. This explains some of the names associated with these proteins isolated from the placenta, that is, chorionic somatomammotropin or chorionic mammotropin. Although PL is implicated in preparation for the mammary gland for lactation, stimulation of steroidogenesis, fetal growth, and alteration of maternal metabolism, direct evidence for effects in cattle are lacking. The best evidence for a role Pl in mammogenesis is in rodents and in sheep and goats.
In contrast with a role for Prl in mammogenesis, there is no doubt about the importance of Prl in lactogenesis. Some of the best evidence for the importance of increased periparturient secretion of Prl in stage II lactogenesis has come from experiments in which the administration of a dopamine agonist was used to inhibit Prl secretion and correspondingly impair lactation. In ruminants where postpartum milking
continues, administration of the dopamine agonist cz-bromoergocryptine (CB154) reduced basal prolactin concentration about 80% and prevented the usual periparturient rise as well as the milking-induced Prl rise during the first week postpartum. Milk production was reduced 45% during the first 10 days postpartum. Lost milk production was associated with reduced synthesis of Oc-Iactalbumin, lactose, and fatty acids, as well as impaired structural differentiation of the mammary secretory cells. Selected effects are summarized in Table 18.6. Cows treated with exogenous Prl in addition to the agonist (to replace the periparturient surge in prolactin) showed no loss of milk production or negative effects on milk component biosynthesis or alveolar cell differentiation. The effect of Prl suppression and replacement during the periparturient period on milk production in multiparous cows is shown in Figure 18.9. Clearly, Prl is important in mammary cell differentiation and lactogenesis (Akers et al., 1981a).
Table 18.6. Changes in lactose synthesis, Oidactalbumin, RNA, and DNA in mammary gland of cows before and after parturition and with treatment with CB154 to suppress Prl secretion.
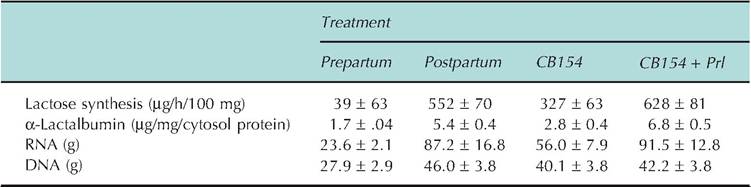
Adapted from Akers et al. (1981b). Control animals were euthanized 10 days before or after parturition. CB1 54 was administered for 12 days prior to expected parturition through parturition. Animals given Prl were infused continuously for 6 days immediately before parturition.
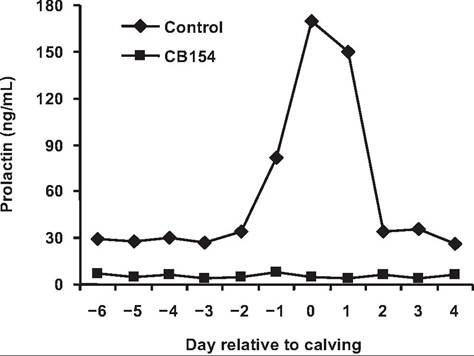
Fig. 18.9. Serum Prl in cows given CB154. Cows were administered daily injections Ofergrocryptine (CB154) during the periparturient period. Both basal concentrations and the surge in Prl, which accompanies parturition, were blocked. Data adapted from Akers et al. (1981 a).
Aside from circumstantial evidence related to changes in circulating hormone concentrations, there is a marked increase in numbers of mammary cell receptors for Prl, IGF-I, and cortisol during late gestation. Progesterone receptor concentration is also correspondingly reduced with the onset of lactation. Thus, simultaneous, coordinated changes in circulating hormones and receptors serve to regulate the timing of lactogenesis. It is believed that high concentration of progesterone during most of gestation acts to inhibit the onset of lactation. Near the time of parturition, progesterone concentrations begin to wane and estrogen concentrations increase. Removal of the negative effects of progesterone along with positive effects of Prl and glucocorticoids sets the stage for the onset of copious milk production (stage II of lactogenesis).
Data from culture experiments also support a role of these hormones in lactogenesis. For example, additions of estradiol or cortisol markedly enhance Prl-induced
secretion of Oc-Iactalbumin (a specific milk protein) by mammary explants taken from pregnant cows. Mammary explants from estrogen-primed or pregnant mice also require insulin, cortisol, and Prl for the accumulation of casein and Oc-Iactalbumin. However, some caution is advised with wholesale extrapolation of results from culture experiments to the intact animal as well as uncritical extrapolation between species. For example, induction of the various milk proteins in culture does not necessarily reflect the timing of events in vivo. Neither do existing culture methods allow consistent synthesis and secretion of milk lipids. Finally, hormone concentrations employed may not accurately reflect the situation at the level of the mammary cell in the animal. Culture systems by their very nature represent relatively uncomplicated regulation compared with the intact animal. Nonetheless, results shown in Figure 18.10 illustrate the positive effects of Prl, cortisol, and estradiol on synthesis and secretion of Oc-Iactalbumin by explants of bovine mammary tissue.
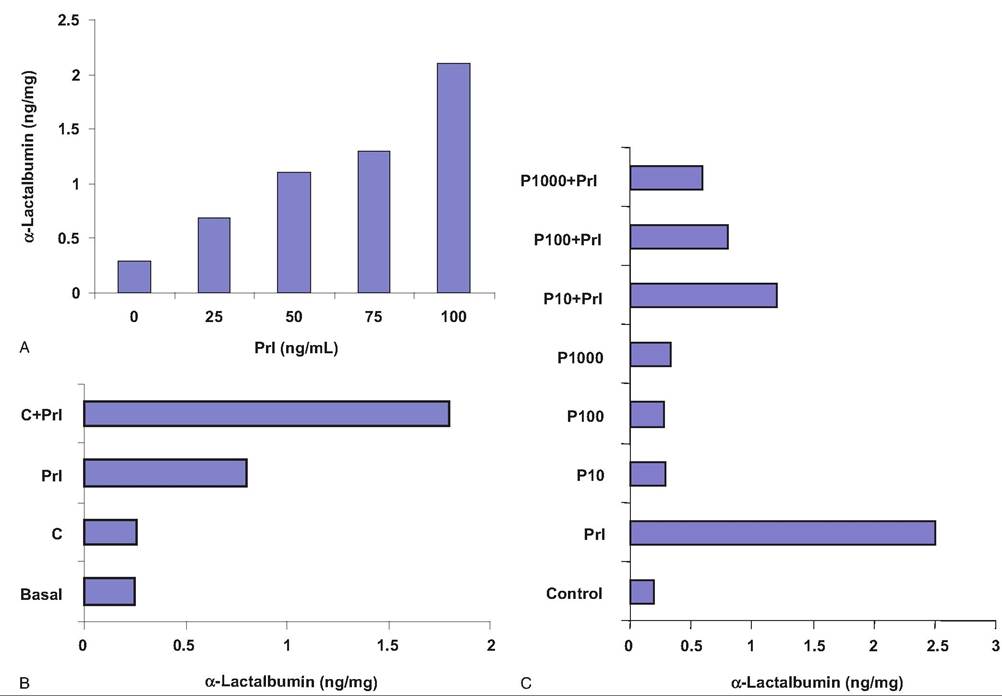
Fig. 18.10. Effect of lactogenic hormones on secretion of α-lactalbumin. Explants of bovine mammary tissue from multiparous, nonlactating, pregnant cows were incubated with various combinations of hormones. Panel A demonstrates a concentration-dependent increase in Oi-Iactalbumin in response to addition of bovine Prl. Panel B shoes the additive effect of cortisol on Prkinduced Oi-Iactalbumin secretion. Panel C demonstrates a concentration-dependent inhibitory effect of progesterone (P) on Prl stimulation of Oi-Iactalbumin secretion. Data adapted from Goodman et al. (1983).
Application of molecular techniques to mammary gland biology has solidified knowledge that Prl and glucocorticoids are the primary stimulators of mammary cell differentiation. Both Prl and glucocorticoid response elements are found within the promoter regions of the genes for several mammary-specific milk proteins. Induction of both mRNA and specific milk proteins in the presence of Prl or glucocorticoids in isolated mammary epithelial cells confirms the importance of these hormones in lactogenesis and provides details for mechanisms of action of these hormones in control of milk protein gene expression (Rosen et al., 1999; Akers, 2006).
The term galactopoiesis was originally coined to describe the enhancement of an established lactation. With this strict definition, only exogenous GH (bST) and thyroid hormone are undisputed galactopoietic hormones in dairy animals. Responses also suggest that these hormones are endogenously rate limiting. However, more generally, galactopoiesis can also be described as the maintenance of lactation. In this sense, a larger number of hormones and growth factors are candidates as galactopoietic agents. Continued secretion of galactopoietic hormones, growth factors, and regular milk removal are essential for regulation and maintenance of lactation following lactogenesis.
The pituitary gland and its hormones are essential integrators of the endocrine regulation of milk secretion. This is dramatically shown by the loss of milk production in hypophysectomized lactating goats. However, milk yield can be fully restored to prehy- pophysectomy levels by the combined administration of Prl, GH, glucocorticoids, and triiodothyronine (T3) see Figure 18.11. Although species differences exist, endocrine organ ablation/replacement studies show
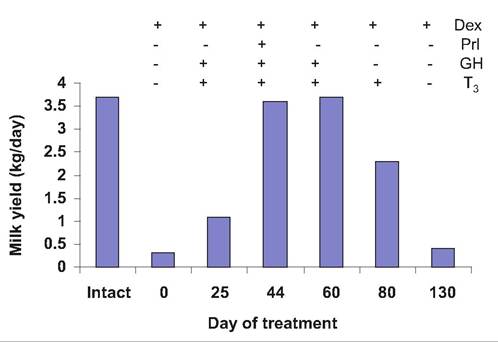
Fig. 18.11. Milk yield after Hypophysectomy. A lactating goat was hypophysectomized and administered hormone replacement. Milk production averaged 3.7kg per day prior to surgery (intact). After treatment for 2 months with dexamethasone (Dex; synthetic glucocorticoid) yields were as indicated at time zero. Milk production is depicted with addition or removal of hormone treatment as indicated. Adapted from Cowie (1969).
that Prl, GH, glucocorticoids, and thyroid hormones are typically required for the full maintenance of lactation (Topper and Freeman, 1980). This does not mean that additional hormones and growth factors (humoral and local, identified and unidentified) might not also be important or mediate effects of these hormones.
Secretion of oxytocin at milking or suckling is necessary in most species for efficient removal of accumulated milk and in some species is essential to obtain any milk at all. The importance of regular milk removal to prevent mammary involution has been known for many years but it has also long been hypothesized that the secretion of galactopoietic hormones with milking or sucking was also important. Secretion of Prl and oxytocin, as well as related secretions of norepinephrine or epinephrine, which can impact the secretion (or action) of oxytocin, are most often associated with hormone secretion at milking and effects on milk synthesis. However, glucocorticoids are also secreted in response to milking or suckling, as is GH in some species.