Neurophysiology
The neuron is a highly excitable cell that is able to respond to and transduce a stimulus into an electrical potential. Like an electrical wire, an axon carries a signal along its length.
Whereas in an electrical wire the signal involves the flow of electrons along the length of the wire, nerve cells are poor conductors of electricity over a long length. Instead of the movement of electrons, neurons rely on the flow of charged ions to propagate the signal. The nerve impulse, or action potential, can travel over long distances without diminishing.The resting membrane potential
Nerve cells and muscle cells have an excitable membrane. When such a cell is not generating an impulse, it is said to be at rest. When at rest, the cytosol has a negative electrical charge relative to the outside of the cell (Fig. 8.9). The potential energy generated by the separation of charge across the membrane is called voltage and is measured in volts or millivolts (IV = IOOOmV). By placing an electrode both inside and outside the cell, a voltage can be measured between these two sites. That voltage is called the potential difference, or potential. The difference in electrical charge across the membrane of a cell at rest is called the resting membrane potential, and it is typically about -60 to -70 mV.
The inside and outside of the cell is separated by a membrane that acts as a Semipermeable membrane. The membrane potential is caused by the unequal distribution of ions across the membrane. For the four
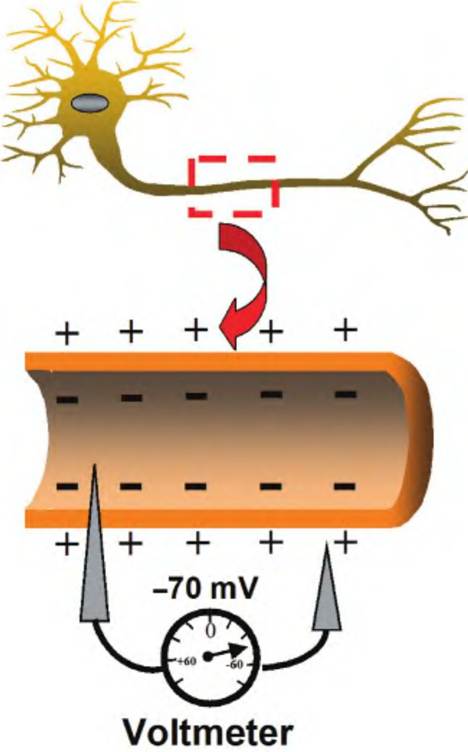
Fig. 8.9. Resting membrane potential. When a voltmeter is used to compare the voltage difference between the inside and outside of a neuron membrane, a membrane potential Ofapproximately -70mV is typically measured.
most important ions with regard to membrane potential, the concentration of Na+ and CF is higher outside the cell, whereas the concentration of K+ and organic anions is higher inside the cell.
The organic anions consist mostly of proteins and amino acids. These differences in ion concentrations establish chemical gradients.The lipid bilayer of the cell membrane acts as an insulator between the interior and exterior of the cell. Ions cannot cross the membrane except by way of ion channels. Within the membrane there are passive ion channels, sometimes called leak channels, which remain open. These passive channels are most permeable to K+ and chloride ions, relatively less permeable to Na+, and impermeable to proteins. Since K+ is in higher concentrations inside the cell, it tends to diffuse outwardly, down its concentration gradient. The movement of ions to eliminate the potential difference is called a current. How much the membrane restricts the movement of ions is a measure of its resistance. As K+ moves outward, an excess of negatively charged anions remain inside the cell, thus establishing an electrochemical gradient. Eventually, the movement of K+ outward reaches equilibrium because the outwardly directed chemical gradient is opposed by the inwardly directed electrical gradient. This point is called the ionic equilibrium potential, or equilibrium potential, and is represented by the symbol Eion.
The equilibrium potential of an ion can be calculated using the Nernst equation:
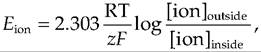
where Eion equals ionic equilibrium potential, R equals the gas constant, T equals absolute temperature, z equals charge on the ion, F equals Faraday's constant, and [ion] is the concentration of the ion either inside or outside the cell. At body temperature, the Nernst equation for the monovalent ions K+, Na+, and CF can be simplified to
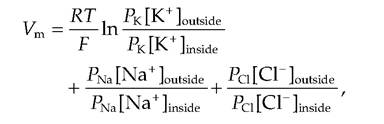
where Vm is the membrane potential and P is the permeability of the respective ion.
The relative permeability of the ions is Pk-Pns-Pci = 1.0:0.04:0.45. Note that in the Goldman equation, the concentrations of CV inside and outside the membrane are reversed compared to Na+ and K+ since CC is an anion.At rest, the flow of ions is not static but is in equilibrium such that Na+ is moving into the cell while K+ is moving out (Fig. 8.10). This movement is countered by an active Na+-K+ ATPase. This active transport system pumps three molecules of Na+ out of the cell for every two molecules of K+ pumped back in. This helps maintain the unequal distribution of ions across the cell membrane (Box 8.1).
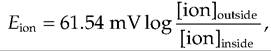
whereas for Ca++, a divalent cation, the equation would be
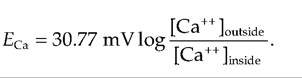
Eion is very important for understanding the effect an ion has on the membrane potential (Table 8.2).
If the membrane were permeable only to K+, the membrane potential would be approximately -80 mV. At this point, the chemical gradient for K+ would equal the electrochemical gradient. However, the membrane is permeable to ions other than K+. The Nernst equation does not take into consideration the permeability of multiple ions, or that the relative permeability of the various ions is different. If the permeabilities of the ions are known, the membrane potential can be calculated using the Goldman equation:
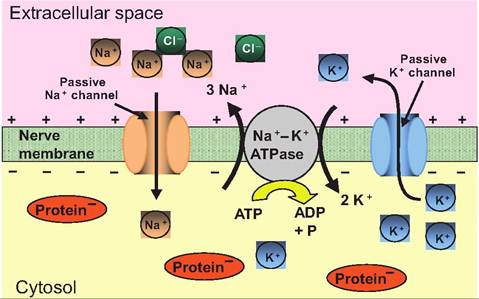
Fig. 8.10. The basis of the resting membrane potential. In a nerve at rest, there is an unequal distribution of ions across the cell membrane. The concentration of Na+ and CF is higher outside the cell, while that of K+ and anions is higher inside the cell.
Moving down their concentration gradient, Na+ passively moves into the cell while K+ moves out of the cell. A Na+-K+ ATPase then actively pumps three molecules of Na+ out of the cell for every two molecules of K+ pumped in while cleaving ATP to ADP plus inorganic phosphate (P).Table 8.2. Ion concentrations and equilibrium potentials in neurons.
| Ion | Concentration outside (mM) | Concentration inside (mM) | Ratio outside:inside | f⅛n (mV) |
| K+ | 5 | 100 | 1:20 | -80 |
| Na+ | 150 | 15 | 10:1 | 62 |
| Ca | 2 | 0.0002 | 10,000:1 | 123 |
| CF | 150 | 13 | 11.5:1 | -65 |
Box 8.1 Lethal injection
Recalling that K+ concentrations are higher inside the cell than outside the cell, imagine what will happen if the extracellular concentration of K+ is suddenly increased by an injection of K+. Using the Nernst equation, it can be shown that an increase in [K+]outside depolarizes the cell, thus eliminating the resting membrane potential. Therefore, an intravenous injection of K+ will cause the heart to stop beating, thus causing death. This, in fact, is how Dr. Jack Kevorkian has helped patients commit suicide by allowing them to switch their intravenous infusion to an anesthetic and potassium solution.
Membrane channels
The neuron is excitable and thus is able to generate and transmit an impulse. A nerve impulse is a change in the resting membrane potential. It is generated in a rather complex process. We have already discussed passive membrane channels that allow ions to move through the membrane of a resting neuron. In addition, there are also active channels, sometimes called gated channels, in the cell membrane.
The gated channels open or close in response to stimuli. There are three classes of gated channels: (1) chemically gated, (2) voltage-gated, (3) mechanically gated.Chemically gated channels
Chemically gated channels, sometimes called neurotransmitter-gated channels, are located on the postsynaptic membrane. These channels are found most abundantly on the dendrites and cell body, and they respond to a neurotransmitter binding to its receptor. The binding of a neurotransmitter will generally cause these channels to either open or close. However, a high concentration of neurotransmitter can cause these channels to become inactivated.
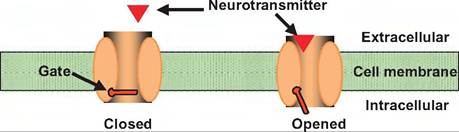
Chemically gated channels can be either directly gated or indirectly gated (Fig. 8.11). Directly gated channels consist of a single macromolecule in which part of the molecule serves as an extracellular domain forming a receptor for a neurotransmitter, while the second part of the molecule consists of a membranespanning domain, which forms an ion channel. Such receptors are sometimes called ionotropic receptors. Binding of the neurotransmitter to the receptor portion of the molecule causes a conformational change that results in opening the channel.
Ionotropic receptors are relatively large and contain multisubunits that form an ion channel through the
1. Directly gated (ionotropic)
2. Indirectly gated (metabotropic)
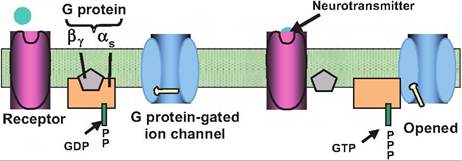
Fig. 8.11. Indirectly and directly gated channel. Neurotransmitters can activate either directly or indirectly gated channels. (1) In directly gated, or ionotropic, channels, the neurotransmitter receptor and ion channel are the same protein. When the neurotransmitter binds to the receptor, it causes a conformation change that results in either an opening or closing of the ion channel. (2) In indirectly gated, or metabotropic, channels, the neurotransmitter binds to a receptor that is distinct from the ion channel.
Binding of the neurotransmitter to the receptor activates a G protein. The G protein can either act directly on a G proteingated ion channel or affect the action of an enzyme that makes a second messenger. GTP, guanosine-5'-triphosphate; GDP, guanosine diphosphate.cell membrane. Binding of a neurotransmitter to such a receptor causes a conformational change that opens the channel permitting ions to flow down their electrochemical gradients. Ionotropic receptors act in submilliseconds.
Indirectly gated channels, often called metabotropic receptors, consist of macromolecules that are distinct from the ion channels they affect. Binding of the neurotransmitter to these channels stimulates or inhibits the production of second messengers such as cAMP, inositol triphosphate, or diacylglycerol. The second messengers then activate protein kinases that are enzymes that phosphorylate other proteins such as ion channels. This can result in opening or closing of ion channels, thus altering the membrane potential.
Metabotropic receptors are composed of a single polypeptide. In contrast to ionotropic receptors, instead of opening an ion channel in response to binding of a neurotransmitter, metabotropic receptors instead normally activate GTP-binding proteins (G proteins). Such receptors generally display slower onset and longer duration than ionotropic receptors.
Voltage-gated channels
Voltage-gated channels are found on those membranes that generate an action potential, including axons and
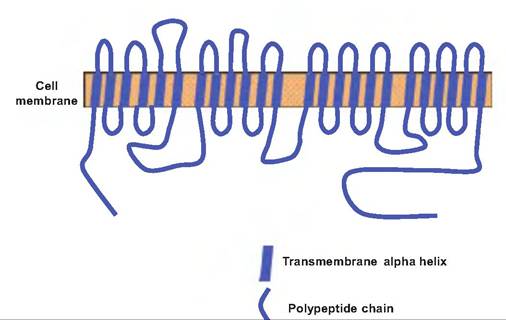
Fig. 8.12. Voltage-gated sodium channel. The voltage-gated sodium channel consists of four groups of homologous domains of similar sequences of amino acids, with each domain being composed of six transmembrane segments. Each transmembrane segment consists of nonpolar amino acids coiled into an alpha helix. The peptide chains connecting the transmembrane domains consist of polar amino acids.
the Sarcolemma of skeletal and cardiac muscle. These channels open or close in response to changes in membrane potential. More precisely, when the membrane potential reaches a critical level called the threshold, voltage-gated channels open. The threshold is normally between -55 and -50 mV.
For our discussion, the most important such channels are those for sodium, potassium, and calcium. The voltage-gated sodium channel is created by a long polypeptide having four distinct domains, each consisting of six transmembrane segments (Fig. 8.12). As the membrane potential becomes more positive due to stimulation, positively charged amino acids lining the pore formed by the channel move away from the center, thus opening the channel. The voltage-gated sodium channel is unique in that it operates in three stages. When stimulated by depolarization, the channel opens rapidly. It remains open for about 1 ms and then becomes inactivated. It cannot be opened again until the membrane potential returns to a negative value below or near the value that originally caused the channel to open. These channels can be blocked by the neurotoxin tetrodotoxin, which is found in the ovaries of the puffer fish (see Box 8.2).
Voltage-gated potassium channels do not open immediately upon stimulation. In addition to having a delayed opening, they also do not become inactivated after stimulation. The voltage-gated calcium channels are similar to the other voltage-gated channels, and will be discussed further with regard to neurotransmitter release.
Box 8.2 Voltage-gated channels: neurotoxins and anesthetics
The puffer fish is considered a delicacy in Japan. When prepared properly, its consumption results in numbness around the mouth. It is prepared by chefs who are licensed by the government for preparation of this fish. Yet each year, dozens of people die from eating this fish. The ovaries of the puffer fish contain a tetrodotoxin, a compound that specifically blocks voltage-gated sodium channels. Blockage of these channels results in the inability of neurons to generate an action potential.
Dinoflagellates of the genus Gonyaulax produce a neurotoxin called saxitoxin that similarly blocks voltage-gated sodium channels. Dinoflagellates can contaminate shellfish such as clams and mussels. Occasionally, there is a sudden increase in the level of dinoflagellates resulting in a "red tide" that causes contamination resulting in occasional human deaths from consumption of these seafoods.
Local anesthetics such as lidocaine also work by blocking voltage-gated sodium channels. Lidocaine binds to the S6 alpha helix of domain IV of the channel, thus blocking the flow of Na+ into the neuron. These anesthetics affect smaller axons before affecting larger axons. Fortunately, it is the smaller axons that carry pain signals.
Mechanically gated channels
These channels open and close in response to physical distortion of the cell membrane. They are found in sensory neurons such as those that respond to touch, pressure, or vibration.
Postsynaptic potentials
At rest, only the passive channels are open, while the neurotransmitter-gated channels are closed. When a neurotransmitter binds to a postsynaptic receptor, it activates a neurotransmitter-gated, or chemically gated, channel that causes a change in the membrane potential. This change in potential is called a postsynaptic potential since it occurs in the postsynaptic cell. Postsynaptic potentials are also graded because their magnitude depends on both the amount and the duration of action of the neurotransmitter.
If the neurotransmitter is excitatory, it generates an excitatory postsynaptic potential (EPSP). During an EPSP, the membrane potential moves toward 0 mV, or becomes depolarized. If the neurotransmitter is inhibitory, it generates an inhibitory postsynaptic potential (IPSP). An IPSP results in the membrane potential becoming more negative, or hyperpolarized. The activation of mechanically gated channels can also cause the generation of an EPSP or IPSP
EPSP
The binding of an excitatory neurotransmitter to a postsynaptic receptor causes channels to open, which allow both sodium and potassium to simultaneously move down their concentration gradient. However, sodium moves inward more rapidly than potassium moves outward due to the electrochemical gradient that exists. Remember that the inside of the cell is negatively charged and thus attracts positively charged molecules. The net result of this ion movement is an increase in intracellular sodium, resulting in depolarization.
The EPSPs are generated in the dendrites and cell bodies. Dendrites and cell bodies do not have voltagegated channels, which are necessary for the generation of an action potential. An EPSP is not the same as an action potential but instead is graded.
IPSP
The binding of an inhibitory neurotransmitter to its receptor causes the cell membrane to become hyperpolarized. Inhibitory neurotransmitters generally cause the opening of transmitter-gated potassium channels, chloride channels, or both. Opening potassium channels allows potassium to move out of the cell, thus making the interior of the cell more negative. Conversely, opening chloride channels allows chloride to move into the cell that also makes the cell more negative.
Generation of an action potential
Neurons are able to generate and propagate an electrical impulse, called an action potential, along their length. The action potential is a stereotypic depolarization and repolarization of the membrane. It is an all-or-none response which means that within a given cell, the response always looks identical both in amplitude and duration, independent of the stimulus. The action potential is generated at one site in the neuron, the axon hillox, and is then propagated down the length of the axon:
Step 1—Resting state. At rest, the voltage-gated channels are closed, and there is only passive movement of ions across the cell membrane.
Step 2—Depolarization. When a neuron receives stimuli, the chemically or mechanically gated
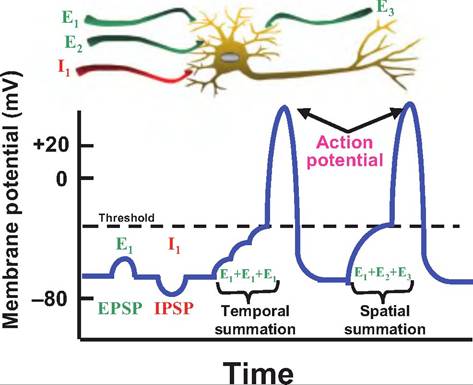
Fig. 8.13. Summation. In response to activation of a chemically or mechanically gated channel, neurons can develop postsynaptic potentials, which can be either excitatory (EPSP) or inhibitory (IPSP). These postsynaptic potentials summate algebraically. EPSPs result in depolarization, whereas IPSPs cause hyperpolarization. Ifa neuron receives stimuli from a single neuron in rapid succession, the postsynaptic potentials can cause temporal summation, whereas if a neuron receives input from multiple locations at the same time, it causes spatial summation.
channels respond. This results in the production of postsynaptic potentials. These are graded potentials, and they can summate (Fig. 8.13). For example, if a neuron generates a volley of EPSPs in rapid succession, they will summate, or add up algebraically, over time, which is called temporal summation. If a neuron receives many stimuli simultaneously from various neurons synapsing on its surface, the postsynaptic potentials summate via spatial summation, meaning that it is occurring across space rather than time. A neuron typically receives both inhibitory and excitatory input that is summated. The postsynaptic potentials are generated in the dendrites and cell's body, and they travel to the axon hillox.
Step 3—Generation of the action potential. If summation brings the membrane to threshold, the voltagegated channels are activated (Fig. 8.14). Upon reaching threshold, the voltage-gated Na+ channels open, increasing the permeability to Na+ a thousandfold and allowing Na+ to rapidly move inward. This causes the membrane potential to move toward the equilibrium potential for Na+ (ENa = +62mV). This results in the upstroke, or rising phase, of the action potential. At the peak of the action potential, the voltage-gated Na+ channels become inactivated. Therefore, Na+ can no longer move inward. However, at this time, the
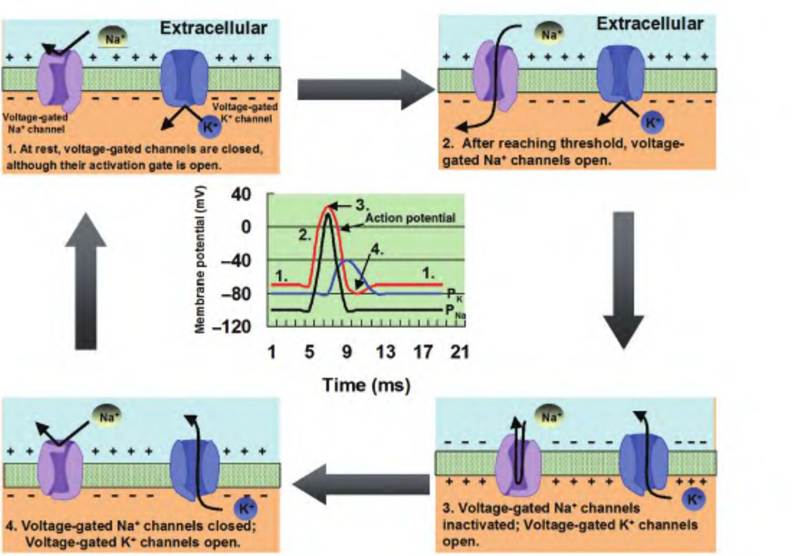
Fig. 8.14. Generation of an action potential. (1) At rest, the voltage-gated Na+ and K+ channels are closed. (2) When the neuron is depolarized to threshold by the summation of postsynaptic potentials, the voltage-gated Na+ channels open. The movement of Na+ into the cell results in the upstroke of the action potential as the membrane potential moves toward the equilibrium potential for Na+. (3) At the peak of the action potential, the voltage-gated Na+ channels become inactivated, while the voltage-gated K+ channels are opening. This results in the downstroke of the action potential as the membrane potential moves toward the equilibrium potential for K+. (4) If the voltage-gated K+ channels remain open for an extended period of time, a hyperpolarizing afterpotential can develop. Finally, the voltage-gated K+ channels close, and the membrane potential returns to rest.
voltage-gated potassium channels are opening, thus allowing K+ to move outward. The outward movement of K+ causes the repolarization phase, also called the downstroke or falling phase, of the action potential in which the membrane potential moves back toward its resting membrane potential.
At the end of the repolarization phase, if the voltagegated potassium channels are slow to close, K+ continues to move outward. This results in a hyperpolarizing afterpotential, or undershoot, in which the membrane potential becomes hyperpolarized relative to the original resting membrane potential before returning to normal.
At the peak of the action potential, the voltagegated sodium channels become inactivated. This creates a brief time period called the absolute refractory period during which no stimuli, no matter how large, can generate an action potential. This is followed by another brief period of time, called the relative refractory period, during which only a greater than normal stimuli can initiate an action potential. A summary of the generation of the action potential is shown in Figure 8.15.
Propagation of the action potential
Once the action potential is generated, it must be carried along the length of the axon. During the generation of the action potential, there is a reversal in membrane potential induced by the movement of Na+ inward, causing the interior of the membrane to temporarily become positive while the outside becomes negative. After this happens, the positively charged ions now inside the membrane move laterally since they are attracted by the negatively charged ions lining the inside of the membrane. Conversely, the positively charged ions found on the outside of the membrane migrate toward the new sink of negatively charged ions created by the reversal in membrane potential, thus completing the circuit.
The local flow of current caused by the opening of the voltage-gated Na+ channels results in a series of electrotonic changes (Fig. 8.16). As the positive ions on the inside of the membrane flow to an area of negative ions, this depolarizes that area and thus causes the voltage-gated Na+ channels in that area to open. A positive feedback system is initiated that allows the action potential to be propagated down the length of the axon without diminishing in amplitude. The
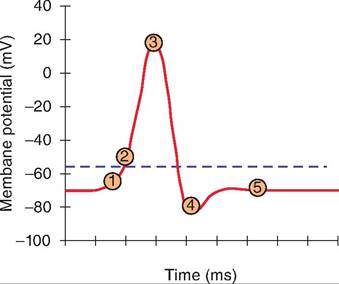
Step 1: Depolarization to threshold— Postsynaptic potentials summate temporally or spatially to reach threshold.
Step 2: Opening voltage-gated Na+ channels—Voltage-gated sodium channels open, allowing Na+to rush inward and membrane potential to move toward ENa+.
Step 3: Inactivation of voltage-gated Na+ channels, activation of voltage-gated K+ channels—At the peak of the action potential, the voltage-gated sodium channels become inactivated. This results in the absolute refractory period during which time another action potential cannot be generated. This is
Immediatelyfollowed by the relative refractory period when a greater than normal stimuli can again open the voltage-gated Na+ channels. At the peak of the action potential, the voltage-gated K+ channels are opening, allowing K+ to flow outward and the membrane potential to move toward Ek-
Step 4: After potential—If the voltage-gated K+ channels are sluggish in closing, then the membrane potential will move closer to the Eκ+ and will drop below the original resting membrane potential.
Step 5: Resting membrane potential—The cell returns to the original resting membrane potential.
Fig. 8.15. Summary of an action potential.
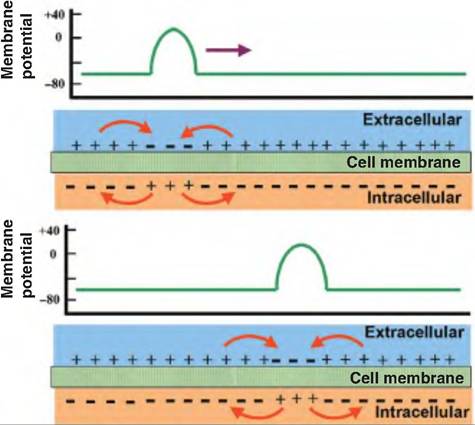
Fig. 8.16. Propagation of an action potential. Local current flows between the site of the action potential and the adjacent inactive areas. This flow of current triggers an action potential in the adjacent area while returning the original site to its resting membrane potential. In this manner, the action potential is able to propagate down the axon.
process is self-propagating. As the action potential migrates down the axon, the voltage-gated Na+ channels in the area it just moved from are absolutely refractory. Therefore, the action potential cannot go backward through that area since the voltage-gated Na+ channels cannot open. Since the action potential originates in the axon hillox in vivo, the action potential moves only from the axon hillox toward the nerve bouton. If the nerve is removed and placed in vitro, and the axon is stimulated in the middle, the action potential can travel in either direction.
Conduction velocity
Conduction velocity refers to the speed with which an action potential is propagated down the length of the axon. Two factors control the conduction velocity: axon diameter and myelin sheath:
• Axon diameter. The larger the axon diameter, the faster the conduction velocity. During an action potential, the cations that enter the cell can move either through the cytoplasm or back through the cell membrane. Both the membrane and the cytoplasm offer resistance. While the cytoplasm offers resistance, the larger the diameter of the axon, the resistance in the cytoplasm becomes less, relative to that of the axon membrane. The farther the current can flow down the axon due to less resistance in the larger diameter axons, the farther the wave of depolarization moves, and thus the faster the speed of conduction.
• Myelin and saltatory conduction. While increasing the diameter of an axon is one strategy to increase conduction velocity, another strategy found in nature is to myelinate the axon. The
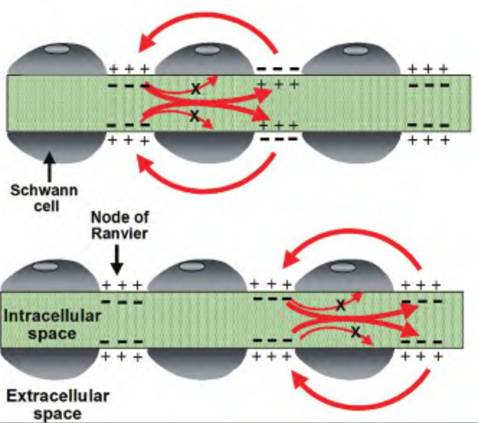
Fig. 8.17. Saltatory conduction. On myelinated axons, the voltage-gated channels are clustered at the node of Ranvier. An action potential propagates along the axon by "jumping" from node to node. Since there are no voltage-gated channels in the internode area, there is no movement of ions nor reversal of membrane potential in these regions.
myelin acts as an insulative sheath enwrapping an axon. There are 0.2- to 2.0-mm breaks in the myelin sheath called nodes of Ranvier. Voltagegated channels are concentrated in the nodes. An action potential is propagated down a myelinated axon by a process called saltatory conduction (from Latin, "to leap"). In essence, the action potential skips along the axon from node to node (Fig∙ 8.17).
Myelinating a neuron also decreases the amount of energy needed for neural transmission. This is because Na+ and K+ move across the axon membrane only at the nodes; therefore, less energy is expended via the Na+-K+ ATPase to restore the Na+ and K+ concentration gradients (Box 8.3).
Box 8.3 Demyelinating diseases
Problems with myelin can result in several disease states. Multiple sclerosis (MS) is a disease involving the myelin sheath of axons in the brain, spinal cord, and optic nerve. Sclerosis is derived from the Greek word for "hardening." In this disease, the myelin sheath in many areas (i.e., multiple) becomes hardened, resulting in a multitude of symptoms such as weakness, lack of coordination, and impaired vision and speech. In Guillain-Barre syndrome, the myelin of peripheral nerves innervating muscle and skin is affected.
Coding of stimulus intensity
If the action potential is an all-or-none response, how does the CNS determine the intensity of a stimulus? How does the brain know whether the animal just touched something warm versus something very hot? The answer lies in the frequency of the action potentials. As a stimulus increases in intensity, a neuron will produce more frequent action potentials. For example, as an area of the skin gets warmer, the frequency of the firing rate of warm receptors will increase. Conversely, as the site gets cooler, the frequency of the firing rate of the warm receptors will decrease, while those of cold receptors will increase.