Synaptic transmission
Electrical transmission
As discussed earlier, electrical synapses are relatively rare. They allow the electrical signal to be propagated from one cell to another with virtually no delay.
Cells are connected via a connexon, and ions can flow from one cell to the other. The electrical signal can be propagated bidirectionally, without modification, across the electrical synapse.Chemical transmission
Far more numerous, chemical synapses are responsible for most communication between neurons and adjacent cells. As shown in Figure 8.5, a chemical synapse is much more complicated. An action potential arriving at the synaptic bouton initiates a series of events:
Step 1—Depolarization of the synaptic bouton. The arrival of the action potential at the nerve ending causes depolarization of this region.
Step 2—Opening voltage-gated calcium channels. Depolarization of the nerve ending allows the voltage-gated calcium channels to open. This allows the influx of calcium from the extracellular space into the synaptic bouton. Calcium causes the synaptic vesicles to bind to the presynaptic membrane. Calcium is then sequestered by the mitochondria or ER, or is actively pumped out of the cell.
Step 3—Exocytosis. Once bound to the presynaptic membrane, the synaptic vesicles release their contents into the synaptic cleft through a process of exocytosis. The vesicles contain quanta, or packages, of neurotransmitter molecules. The neurotransmitter diffuses throughout the synaptic cleft. The amount of neurotransmitter released is dependent on the frequency of impulses reaching the synaptic bouton. The higher the frequency of impulses, the more synaptic vesicles that bind to the presynaptic membrane, and the more neurotransmitter released.
Step 4—Binding of neurotransmitter to postsynaptic membrane. If the neurotransmitter binds to a postsynaptic receptor, it activates the transmitter-gated channels.
This will result in either an IPSP or EPSP in the postsynaptic cell, depending on whether the neurotransmitter is inhibitory or excitatory.Step 5—Inactivation of the neurotransmitter. The neurotransmitter can continue to cause an effect on the postsynaptic cell until it is inactivated. The action of neurotransmitters is short-lived due to rapid methods of inactivation. The methods of inactivation include (1) enzymatic breakdown, (2) reuptake into the neurons that released the neurotransmitter, (3) uptake by glial cells, or (4) diffusion from the site.
During chemical transmission, there is a synaptic delay of 0.2-0.5 ms between the time an impulse reaches the nerve bouton and the initiation of a postsynaptic potential in an adjacent cell. This delay is due to the time it takes for all the events associated with chemical transmission, including (1) opening voltagegated calcium channels, (2) entry of calcium, (3) binding of synaptic vesicles to the presynaptic membrane, (4) exocytosis, (5) diffusion of the neurotransmitter to postsynaptic membrane, and (6) activation of transmitter-gated channels.
Modulation of the synaptic signal
The transmission of a signal across the chemical synapse can be modified both over the short or long term. This is known as synaptic plasticity. It can be controlled by either internal processes within a neuron or by extrinsic processes.
For short-term regulation, the amount of transmitter released at the synapse is dependent on the concentration of presynaptic calcium. Calcium enters the presynaptic terminal through L-type voltage-gated Ca2+ channels. Hyperpolarizing or depolarizing the resting membrane potential can decrease or increase the entry of Ca++ through these channels, thus affecting neurotransmitter release. In addition, another neuron can synapse on the axon terminal of the neuron and also control the amount of neurotransmitter released by that cell. This can cause either presynaptic inhibition or facilitation depending on whether the amount of neurotransmitter released is decreased or increased, respectively (Fig.
8.18).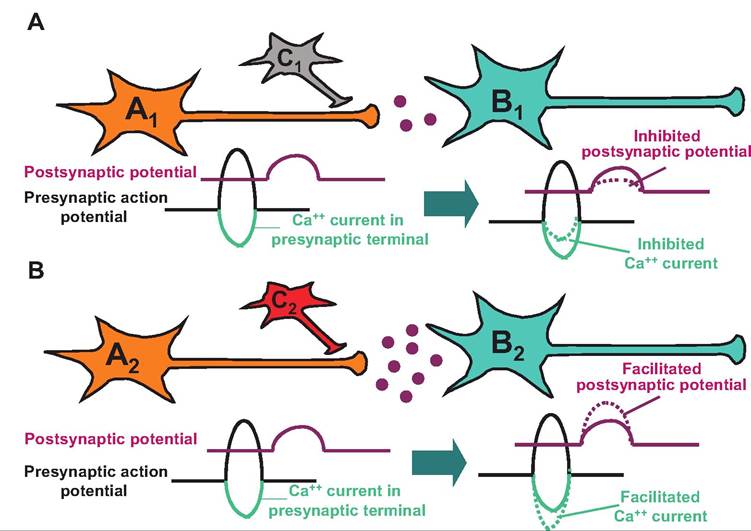
Fig. 8.18. Presynaptic inhibition (A) and presynaptic facilitation (B). (A) An inhibitory neuron (C1) can synapse on the axon terminal of a presynaptic neuron (A1). Release of a neurotransmitter from C1 can Ityperpolarize A1 reducing the influx of Ca into the presynaptic terminal and thereby decreasing the amount of neurotransmitter released from A1, which acts on neuron B1. (B) An excitatory neuron (C2) can synapse on a presynaptic neuron (A2). Release of the excitatory neurotransmitter can enhance the entry of Ca into neuron A2 and thereby increase the release of a neurotransmitter from A2, which acts on neuron B2.
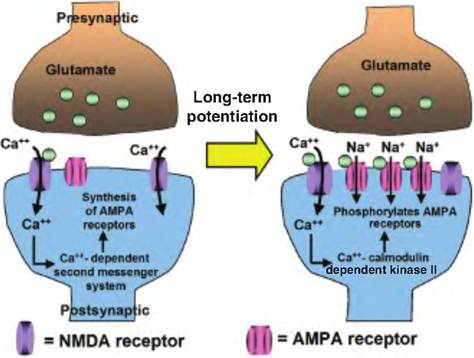
Fig. 8.19. Long-term potentiation. Stimulation Ofpresynaptic glutaminergic neurons causes the release of glutamate. Glutamate binds to postsynaptic glutamate NMDA receptors leading to the entry of Ca into the postsynaptic cell. Ca stimulates a Camdependent second messenger system, resulting in the formation of new AMPA glutamate receptors. Thereafter, stimulation of NMDA receptors allows the entry of Ca, which stimulates Ca - Calmodulin dependent kinase Il to phosphorylate AMPA receptors. The AMPA receptors then allow the entry of Na+, which results in enhanced depolarization of the postsynaptic cell, and thus long-term potentiation.
The neurotransmitter gamma-aminobutyric acid (GABA) has been shown to cause presynaptic inhibition by increasing the presynaptic influx of CB through GABA-gated CB channels. This causes hyperpolarization of the presynaptic neuron that results in less influx of Ca++ into the presynaptic terminal in response to an action potential in that cell. In contrast, serotonin has been shown to activate a cAMP-dependent protein kinase causing phosphorylation and closure of K+ channels.
This results in depolarizing the presynaptic terminal, allowing the entry of Ca++ to persist longer, and thus enhancing the release of the neurotransmitter from the presynaptic neuron.Calcium also appears involved in another type of synaptic plasticity that lasts for a longer period of time called long-term potentiation (Fig. 8.19). Stimulation of the glutamate NMDA receptor, named after the chemical N-methyl D-aspartate, results in an influx of Ca++ into the postsynaptic nerve terminal. Although this does not cause an EPSP, it does cause activation of a Ca++-dependent second messenger pathway, which results in an increase in the number of another type of glutamate receptors called α-amino-3-hydroxy 5-methylisoaxzole-4-propionic acid (AMPA) receptors at the same site. Now, when glutamate stimulates NMDA receptors, thus allowing Ca++ to enter, Ca++ stimulates calcium-calmodulin-dependent protein kinase II, which phosphorylates AMPA receptors. The AMPA receptors then open and allow Na+ to enter, resulting in enhanced formation of EPSPs. Because of the increased number of AMPA receptors, the cell now displays an enhanced sensitivity to glutamate resulting in long-term potentiation.
Neurotransmitters
Neurotransmitters are the means by which signals are carried across the chemical synapse. To be considered a neurotransmitter, a compound must satisfy the following four criteria:
1. It must be synthesized in the neuron.
2. It must be present in the presynaptic terminal and released in amounts sufficient to exert a defined action on the postsynaptic cell.
3. When administered exogenously, it must exactly mimic the action of the endogenously released neurotransmitter.
4. There must be a specific mechanism of inactivation.
It was originally believed that neurons released a single neurotransmitter. This is known as Dale's principle. Although some neurons make and release a single neurotransmitter, it is now known that some neurons release more than one neurotransmitter.
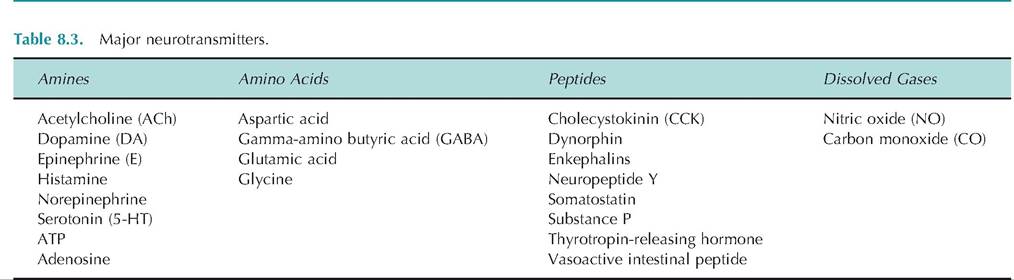
It is not uncommon for a peptide-containing neuron to also release an amino acid or amine-type neurotransmitter. If a cell releases more than one neurotransmitter, it can either be coreleased or released separately in response to various stimulation frequencies.Neurotransmitters are generally grouped into four chemical categories: (1) amino acids, (2) amines, (3) peptides, or (4) dissolved gases (Table 8.3). The term biogenic amines has classically been used to include the catecholamines and serotonin. Occasionally, histamine is also considered a biogenic amine. The term classical neurotransmitters refer to ACh, biogenic amines, and the amino acid neurotransmitters.
Amines
ACh
ACh was the first neurotransmitter discovered. It is the transmitter at the neuromuscular junction. It is therefore synthesized in all motor neurons whose cell bodies are in the spinal cord and brain stem. It is also used in other cholinergic neurons both in the CNS and PNS (Box 8.4).
Synthesized in a one-step enzymatic process within the nerve ending, choline acetyltransferase transfers an acetyl group from acetyl CoA to choline to form ACh (Fig. 8.20). Once synthesized in the cytosol of the
Box 8.4 Acetylcholinesterase (AChE)
Many pharmacological agents have been developed in order to manipulate the synaptic levels of acetylcholine (ACh). AChE inhibitors increase the synaptic levels of ACh. Examples of such inhibitors include various nerve gases, such as sarin, that were developed for chemical warfare and organophosphates, such as parathion, which are used as insecticides. Succinylcholine, which is a competitive neuromuscular blocking agent, is used as an adjunct with anesthetics to increase muscle relaxation during surgery.
cell, ACh is concentrated into the synaptic vesicles by an ACh transporter. Upon stimulation of the cell, ACh is released into the synaptic cleft where it can bind to a cholinergic receptor or be enzymatically degraded by AChE.
AChE is secreted into the synaptic cleft, and its action represents the major route of inactivation for ACh. The action of AChE yields choline and acetic acid. Choline is taken back into the cholinergic axon terminals by a specific choline transporter. Since the availability of choline limits how much ACh can be synthesized, the transport of choline into the presyn- aptic terminal is the rate-limiting step.Catecholamines
The amino acid tyrosine is the precursor to a group of neurotransmitters that all contain the chemical structure called a catechol. Collectively, this group of neurotransmitters is called catecholamines and include dopamine, norepinephrine, and epinephrine. Catecholamines are compounds that contain both a catechol and an amine group (Fig. 8.21). Since norepinephrine (NE) and epinephrine (E) are found in the adrenal medulla, they are sometimes called norepi
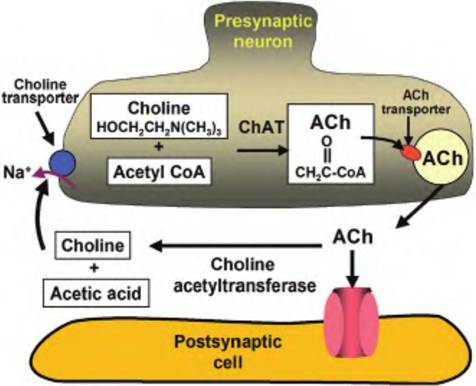
Fig. 8.20. Cholinergic neuron. Choline plus acetyl CoA, acted upon by choline acetyltransferase, combine to form acetylcholine (ACh). ACh is actively pumped into the synaptic vesicles by an ACh transporter. When released from the presynaptic cell, ACh can either bind to a presynaptic receptor or be broken down by choline acetyltransferase into choline and acetic acid. Choline is then actively pumped back into the presynaptic cell by a choline transporter where it can be used to make more ACh.
nephrine and epinephrine, respectively. Catechol- aminergic neurons are found throughout the peripheral and CNS. The role of epinephrine within the CNS is limited since it is found in only two medullary sites, known as the Cl and C2 cell groups.
The first step in the synthesis of catecholamines is catalyzed by tyrosine hydroxylase that converts tyrosine to L-dihydroxyphenyIalanine (L-dopa). This step is under end-product inhibition so that if the levels of catecholamines in the synapse rise, they can inhibit the activity of this enzyme. Furthermore, when catechol- aminergic neurons are stimulated, enzymatic activity of this enzyme increases in response to depletion of
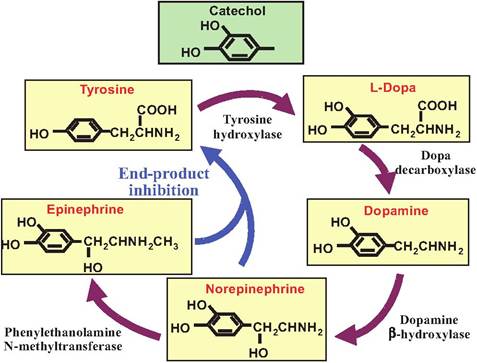
Fig. 8.21. Catecholamine synthesis. Catecholamines are a group of compounds that contain both a catechol and an amine group. Synthesized from tyrosine, the catecholamines include dopamine, norepinephrine, and epinephrine. The rate-limiting enzyme, tyrosine hydroxylase, is under end-product inhibition. Ifa neuron releases only dopamine, it will lack those enzymes and compounds found further along in the biosynthetic pathway.
the catecholamines. The remainder of the biosynthetic pathway is shown in Figure 8.21.
Catecholamines are inactivated at the synapse by selective reuptake into the presynaptic terminal. This reuptake system involves Na+-dependent active transporters. These transporters are subject to pharmacological manipulation. For example, amphetamine and cocaine can block these transporters, thus prolonging the action of the neurotransmitter in the synapse. After reuptake, the neurotransmitters can be enzymatically degraded by monoamine oxidase (MAO), or they can be reloaded into the synaptic vesicles. Another enzyme, catechol-O-methyItransferase, is also found in the synapse and can break down catecholamines.
Indoleamine
The neurotransmitter serotonin, also called 5-HT, is an indoleamine. It is synthesized from the amino acid tryptophan (Fig. 8.22). While in relatively low concentrations in the CNS, 5-HT is involved in mood, emotional behavior, and sleep. Because the synthesis of 5-HT is not under end-product inhibition, the endogenous levels of 5-HT can be increased by dietary supplementation with tryptophan.
Once in the synapse, 5-HT is inactivated by a specific serotonergic reuptake transporter. Like the transporter for catecholamines, this transporter is subject to
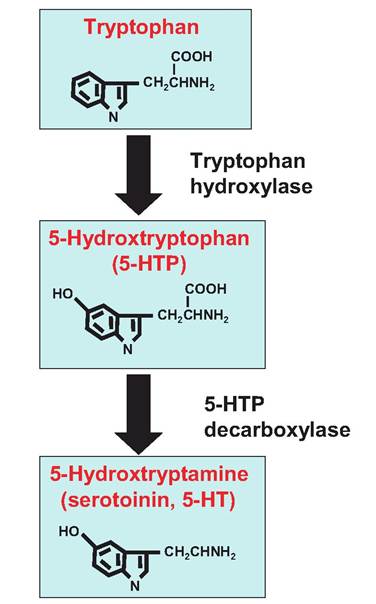
Fig. 8.22. Serotonin synthesis. The indoleamine serotonin, also called 5-hydroxytryptamine, is synthesized from the amino acid tryptophan. The synthesis of serotonin is not under end-product inhibition.
pharmacological manipulation. One very successful class of antidepressants is known as specific serotonergic reuptake inhibitors (SSRIs), which include Prozac®. Once inside the cell, 5-HT is degraded by MAO.
Adenosine
There is growing evidence that ATP, and its derivatives such as adenine, can act as neurotransmitters. Adenine and guanine, and their derivatives, are called purines. These compounds act as purinergic receptors. Adenosine acts to dampen sympathetic function following intense sympathetic activity, thus reducing further release of norepinephrine and ATP, which are coreleased from sympathetic fibers.
Amino acids
Amino acids are ubiquitous within the body, and some appear to act as neurotransmitters. The amino acids glutamate and aspartate act as excitatory
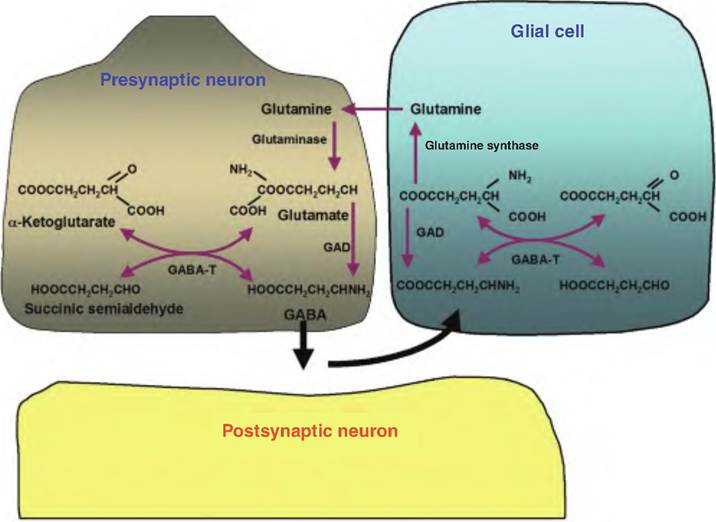
Fig. 8.23. Gamma amino butyric acid synthesis (GABA). GABA is ultimately synthesized from glucose metabolism in which a-ketoglutarate formed in the Krebs cycle is transaminated to glutamate catalyzed by the enzyme α-oxo-glutarate transaminase (GABA-T). The amino acid glutamate is decarboxylated by the enzyme glutamic acid decarboxylase (GAD). Upon release, GABA is taken back into the presynaptic neurons or glial cells. Once inside the glial cells, glutamate is formed. Glutamate must be converted to the noncharged glutamine by glutamine synthetase in order to be transported back to the presynaptic neuron where it is deaminated by glutaminase.
neurotransmitters, whereas glycine acts as an inhibitory neurotransmitter in the interneurons of the spinal cord. Although these amino acids are found in all cells, the concentrations are about two to three times higher (-20 mM) in those cells in which they are neurotransmitters. Furthermore, there are transporters within those cells that load the amino acids into vesicles, thus further increasing their concentrations (50mM). These transporters are found only in neurons in which the amino acid is a neurotransmitter.
The neurotransmitter gamma aminobutyric acid (GABA) is synthesized from the amino acid glutamate (Fig. 8.23). However, unlike other amino acids that are found ubiquitously throughout the body, GABA is found only in neurons that use it as a neurotransmitter. GABA serves as the neurotransmitter at most CNS synapses where it acts as an inhibitory neurotransmitter. It is also found in other tissues, including the islet cells of the pancreas and adrenal gland. GABA is inactivated by uptake into the presynaptic terminals and glia by Na+-dependent transporters.
Histamine, in addition to serving a role in the immune system where it is found in mast cells, also acts as a neurotransmitter. It is localized in the hypothalamus. It is synthesized from the amino acid histidine by decarboxylation (Fig. 8.24).
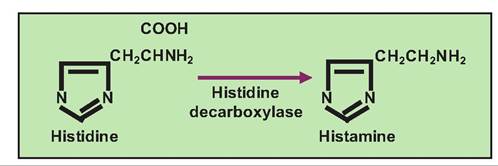
Fig. 8.24. Histamine synthesis. Histamine is synthesized from histidine by the action of histamine decarboxylase.
Peptides
Peptides consist of strings of amino acids connected in amide linkages. Unlike the classical neurotransmitters discussed earlier, which are synthesized in the nerve ending, peptides are synthesized in the cell body and transported to the nerve ending (Fig. 8.25). Peptides are generally synthesized as a prohormone in the rough ER. This prohormone is packaged in secretory vesicles that bud off from the Golgi apparatus. Peptidases then act on the prohormone to produce the active transmitter. In addition to being synthesized differently from classical neurotransmitters, peptides are inactivated by enzymes rather than by reuptake processes.
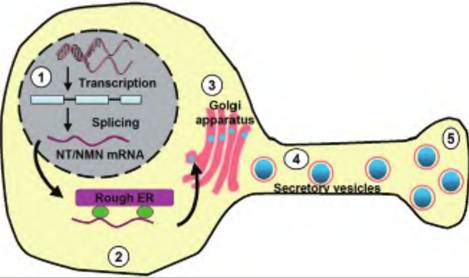
Fig. 8.25. Peptide neurotransmitter synthesis. Peptide neurotransmitters are generally synthesized by transcription in a prohormone form. (1) For example, the 1 70 amino acid prohormone precursor of neurotensin (NT) is encoded by a single gene. (2) The mRNA is translated by the rough endoplasmic reticulum (ER). The prohormone that is made also contains one copy each of neurotensin and neuromedin N (NMN). (3) The prohormone is targeted to the Golgi apparatus where it is packaged into secretory vesicles. (4) The secretory vesicles are transported anterogradely toward the nerve bouton. (5) The peptide transmitter is ready for release by the process of exocytosis.
Whereas the amounts of classical neurotransmitters can be enhanced through increased local synthesis, peptide transmitters require activation of gene expression that takes hours or days. The classical neurotransmitters are generally stored in relatively small vesicles (~50nm), whereas peptides are stored in larger vesicles IOOnm in diameter. In addition, peptide neurotransmitters are typically released at higher neuronal firing rates than are necessary to release the classical neurotransmitters.
Unconventional neurotransmitters
Nitric oxide
Although its mechanism was unknown, for decades, nitroglycerine was used to treat cardiovascular disorder. In the 1980s, it was known that a factor called endothelial-derived relaxation factor was produced by the cells lining blood vessels and caused vasodilation. This factor was shown to be nitric oxide (NO). Nitric oxide is an unconventional neurotransmitter for several reasons: (1) It is a gas; (2) it is not stored; (3) it is not released in an exocytotic manner; and (4) there is no active process that terminates the action of NO. Instead, since NO is small and uncharged, it readily crosses membranes, and due to its short half-life of less than 30 seconds, it decays spontaneously to nitrite.
The synthesis of NO is very simple, involving one enzymatic step in which L-arginine is converted to citrulline and NO (Fig. 8.26). The enzyme that catalyzes this reaction is nitric oxide synthase (NOS). There are three known isoforms of NOS, including
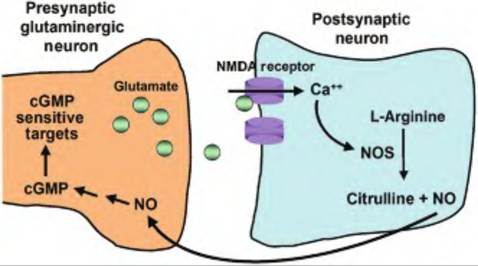
Fig. 8.26. Nitric oxide (NO) containing neuron. Nitric oxide is synthesized from arginine by the action of nitric oxide synthase (NOS) that forms citrulline and NO. Glutamate, acting on NMDA receptors, causes an influx of Ca into the postsynaptic cell. Calcium then induces NOS. Nitric oxide, which is small and membrane-permeable, acts as a retrograde messenger causing the synthesis of cGMP in the presynaptic cell.
macrophage-inducible NOS present in microglia, endothelial NOS found in endothelial cells lining blood vessels, and neuronal NOS.
NO can act as a retrograde messenger, meaning that it is produced in a postsynaptic cell and acts on the presynaptic cell where it activates guanylyl cyclase, resulting in the production of cGMP, which then enhances the release of the neurotransmitter from the presynaptic cell. As such, NO is believed to facilitate long-term potentiation in which the release of a neurotransmitter from a presynaptic cell is increased upon increased firing rate at the synapse.
Carbon monoxide
Carbon monoxide (CO) is a gas that forms in the body by the enzyme heme oxygenase-2, the same enzyme responsible for degrading heme in aging red blood cells. This enzyme is localized in discrete neuronal populations suggesting that CO, like NO, may act as a neurotransmitter. In fact, from using knockout mice (i.e., mice that have a gene removed or inactivated), it appears that CO and NO both function in the relaxation of smooth muscle associated with peristalsis in the gastrointestinal tract. Like NO, CO activates gua- nylyl cyclase.
Neurotransmitter receptors
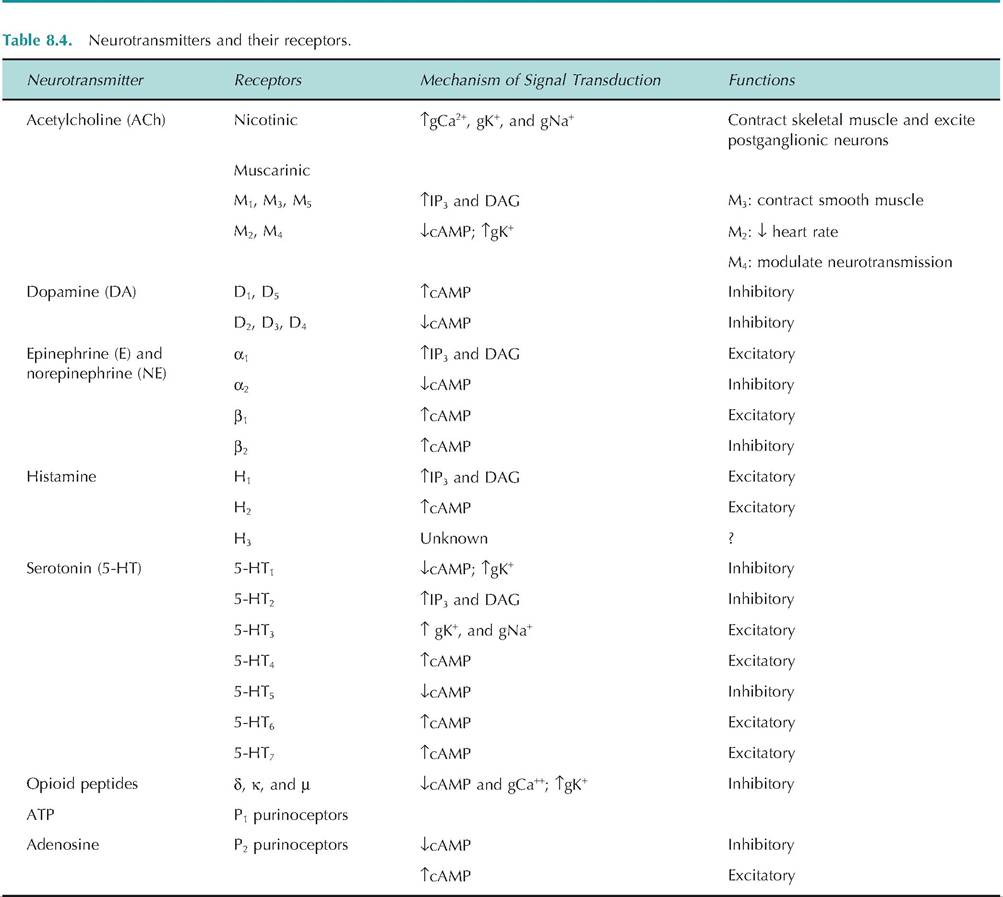
The action of neurotransmitters is dependent on the receptor to which they bind. Neurotransmitters can generally act at multiple receptors, resulting in differential responses. Each of the receptors to which a neurotransmitter binds is called a receptor subtype.
For example, ACh can bind to both nicotinic and muscarinic receptors. Although ACh can bind to both receptor subtypes, the subtypes mediate
DAG, diacyl glycerol; g, conductance; T, increased; X, decreased.
different responses. Nicotinic receptors are found in skeletal muscle where their stimulation results in contraction of skeletal muscle. In contrast, muscarinic receptors are found on postsynaptic cells of the parasympathetic nervous system, and their stimulation can cause contraction of smooth muscle or decreased heart rate.
The same neurotransmitter can cause opposite effects depending on which receptors it stimulates. When norepinephrine binds to β2-adrenergic receptors, it causes vasodilation, whereas binding to α1-adrenergic receptors causes vasoconstriction. Examples of receptors and their effects are shown in Table 8.4.
Neuropharmacology deals with the action of various neurotransmitters and explores drugs that either mimic or block the action of neurotransmitters. A drug that mimics the action of a neurotransmitter at its receptor is called an agonist; a drug that blocks the neurotransmitter is called an antagonist (Table 8.5).
Agonists and antagonists are routinely used in animal and human medicine. They have been developed because they generally have fewer side effects because of their specificity and have longer half-lives than neurotransmitters. Treatments typically involve either stimulating or inhibiting the action of a neurotransmitter. Since atropine can block the action of the parasympathetic nervous system, it is commonly given prior to surgery in order to reduce the production of saliva and thereby reduce the chances of suffocation during surgery. In glaucoma, it is necessary to increase smooth muscle tone in order to increase outflow of aqueous humor. The administration of ACh would be ineffective because of its rapid hydrolysis by AChEs. Instead, a muscarinic agonist, such as pilocarpine, is given.
Table 8.5. Neurotransmitters: agonists and antagonists.
