Overview of male reproductive tract
The reproductive tract of the male is made up of testis, epididymis, spermatic cord, accessory sex glands, and the penis. The testis is critical for production of spermatozoa and testosterone.
The epididymis provides the environment needed for the spermatozoa to mature and storage. The accessory sex glands produce seminal plasma and fluids and the penis the structure needed for copulation.The testes lie outside the abdominal cavity within the scrotum, essentially a purse-like structure that is derived from the skin and connective tissue of the abdominal wall. The testes develop in the abdomen, in a position medial to the embryonic kidney. A grouping of ducts within the testis develops so that they connect to the mesonephric tubules to eventually give rise to the epididymis, ductus deferens, and the vesicular gland. The prostate and bulbourethral glands develop from the embryonic sinus and the penis forms by tabulation and elongation of a tubercle that appears at the orifice of the urogenital sinus. This pattern of development depends on secretions from the fetal testis. Specifically, androgen production stimulates secretion of a glycoprotein called Mullerian-Inhibiting substance, which prevents the female developmental pattern from proceeding. In other words, in the absence of blockage, the female development pattern is normal.
The testes normally migrate in a caudal direct within the abdominal cavity to the inguinal ring. They pass through the enlarged foramen of the genitofemoral nerve (Lumbar; L3 and L4). This process requires the formation of a peritoneal sac that extends through the abdominal wall so that the testis is enclosed by an inguinal ligament of the testis called the gubernaculum testis. The migration is complete once the testes are in the scrotum. If this process fails, the condition is called crytorchidism. If both testes fail to descend, the male is sterile.
This is because the lower temperature of the testes within the scrotum is required for spermatogenesis. As diagrammed in Figure 19.16, the various parts of the male reproductive system have been likened to a manufacturing operation. If successful, the end products are fully functional spermatozoa. This requires hormones (testosterone, FSH, and LH) and secretory products (fluids of the epididymis and seminal plasma) to allow for appropriate movement, transport, and maturation of the spermatozoa.The testes are responsible for the creation of the spermatozoa and they are very efficient. Depending on the species, 1-25 billion spermatozoa are produced per day. Once the cells are produced over a period of several days, they pass thorough the rete tubules and enter the head (caput) and body (corpus) of the epididymis. In these locations the cells are modified so that they become fertile. Cells that enter the tail (cauda) of the epididymis are capable of fertilization if removed and incubated in an appropriate buffer solution. This region of the epididymis serves as a reservoir for spermatozoa prior to ejaculation. With sexual arousal, the spermatozoa in the tail of the epididymis are transported via contractions of the smooth muscle
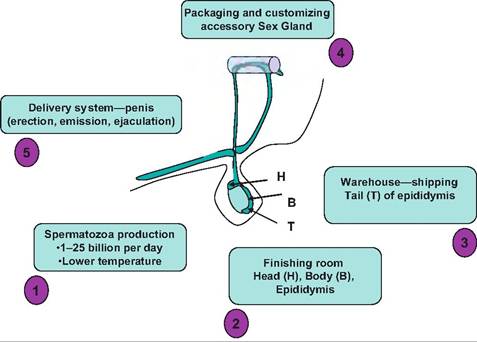
Fig. 19.16. Male reproductive function viewed as a manufacturing process. (1) Production of spermatozoa occurs in the seminiferous tubules of the testes; (2) spermatozoa are transported to the body and head of epididymis; (3) in the tail of the epididymis, spermatozoa become functionally fertile; (4) addition of secretions from the accessory sex gland (seminal vesicles, prostate, etc.) provides seminal plasma; (5) sexual arousal and intromission provide the delivery system to the female reproductive tract.
of the epididymal duct and the ductus deferens to the pelvic urethra. The final packaging involves the addition of fluids produced by the accessory sex glands (ampulla of the ductus deferens, seminal vesicles, prostate gland, and bulbourethral glands).
The non- Cellular component of the semen is called seminal plasma. Some of these basic structural elements are illustrated in Figure 19.17.Spermatogenesis
The functional tissue of an organ is called the parenchyma of the organ. Testicular parenchyma is comprised of the seminiferous tubules and the interstitial cells of Leydig. The testis is surrounded by a tough capsule composed of two elements. The tunica vaginalis (visceral tunic) is closest to the testicular parenchyma and the tunica albuginea, a tougher outer connective tissue layer to the outside. Tissue projections from the capsule penetrate into the testicular tissue to create supporting septa for the seminiferous tubules where they join a central connective tissue band called the mediastinum. When the testis is cut longitudinally it appears that there are elongated islands of parenchymal tissue separated by faint bands of connective tissue. Examples of the gross and microscopic anatomy of the testis are shown in Figure 19.18.
The seminiferous tubules are the key to spermatogenesis. As the images in Figure 19.17 and Figure 19.18
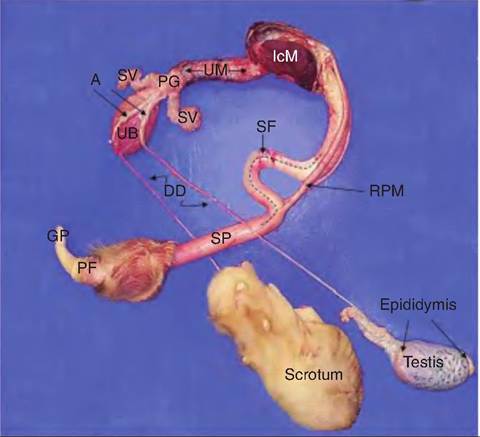
Fig. 19.17. Gross anatomy of the dissected reproductive tract of a bull, A=ampulla of ductus deferens, GP=glans penis, DD=ductus deferens, IcM= Ischiocabernosus muscle, RPM=retractor penis muscle, PG = prostate gland, PF = preputial fold, SF = sigmoid flexure, SP = shaft of penis, SV = seminal vesicles, UB = urinary bladder, UM = urethralis muscle.
suggest, they are very highly convoluted structures with a complex array of developing cells (germinal epithelium) that make up the wall of the tubule. The end of each seminiferous tubule joins with the rete tubules that are continuous with the epididymis. The tubule is surrounded by peritubular cells, connective tissue elements, capillaries, and the Leydig cells. Within the basement membrane the basal compartment contains the Sertoli cells and the developing population of germ cells the spermatogonia.
The cycle of differentiation involves sequential development of the germ cells until the mature spermatozoa are released into the luminal space of the seminiferous tubule. The Sertoli cells are the only somatic cells that are part of the internal structure (inside the basement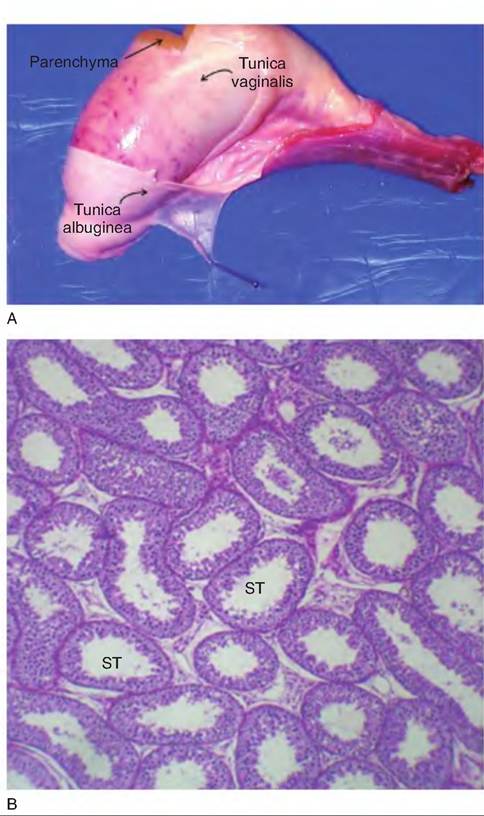
Fig. 19.18. Gross anatomy of the bull testis and corresponding histological appearance of the testicular parenchymal tissue. The capsule surrounding the testis (tunica vaginalis and tunica albuginea) supports the parenchymal tissue (panel A). The striking feature of the testis is the abundance of seminiferous tubules (ST), responsible for the production of the spermatozoa (panel B).
membrane) of the seminiferous tubules. As the spermatogonia develop, their destiny is largely controlled by the Sertoli cells. Each of the Sertoli cells tends or anchors a maximum number of germ cells. This means that the absolute number of spermatozoa produced per day is directly linked with the density of Sertoli cells within the seminiferous tubules. In many ways the Sertoli cells are analogous to the granulosa cells of the ovarian follicles. The layer of Sertoli cells near the basement membrane are linked to one another by tight junctions. This produces a barrier that allows different local environments within the basal region compared with the area near the lumen of the tubule (the adluminal compartment). The basal region houses the spermatogonia and the early spermatocytes. The adluminal region has all the other germ cells, that is, primary and secondary spermatocytes, spermatids, elongated spermatids, and mature spermatozoa. Cells of the seminiferous tubules are illustrated in more detail in Figure 19.19. Because not all the cells in regions of the tubules mature simultaneously, a survey of sections shows varying stages of maturity. Figure 19.20 shows a section of seminiferous tubule with fully formed spermatozoa nearing the point of being released into the lumen; this would be Stage VIII of the development.
This developmental process can be characterized in several ways. A cycle is described as a series of changes in a given region of the tubular epithelium between appearances of the same developmental stage. A stage
is a well-characterized histological-cellular profile of the epithelium that defines a particular period of spermatogenesis. A wave of development defines the sequential order of the stages of spermatogenesis that occur along the length of the tubule. As an example, if you begin at the time when a "crop" of new spermatozoa have been released into the lumen of the tubule, this region of the germinal epithelium is
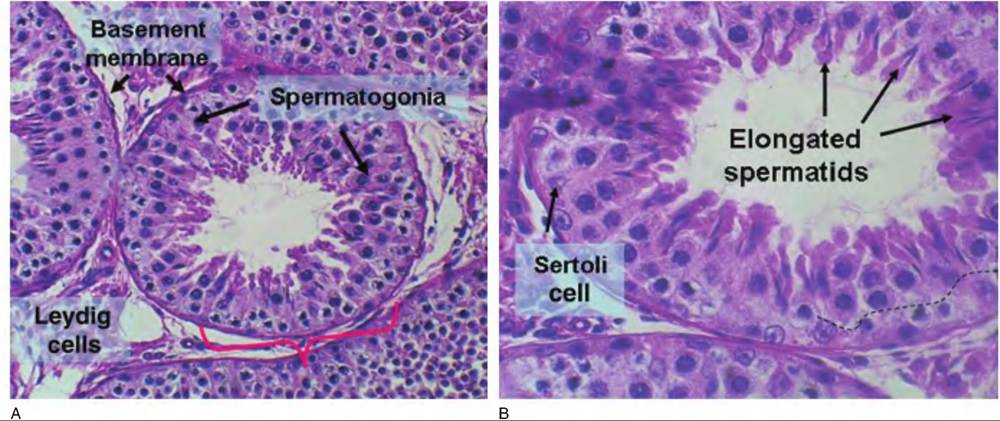
Fig. 19.19. Histology of seminiferous tubules is illustrated. Panel A provides an overview of the seminiferous tubules with their distinct basement membranes (bracket indicates a portion of a single tubule). In the regions between the tubule islands of cells, Leydig cells (lower left corner) are responsive to gonadotropin (FSH and LH) secretion and in turn produce locally high concentrations of testosterone. Testosterone, in addition to its effect on secondary sex characteristics, is also important in differential of the germ cells. Panel B provides higher magnification to illustrate the presence of Sertoli cells and spermatogonia of the basal compartment as well as presence of spermatids and elongated spermatids in the adluminal compartment. The thin dashed line shows the approximate boundary between the basal and adluminal compartments.
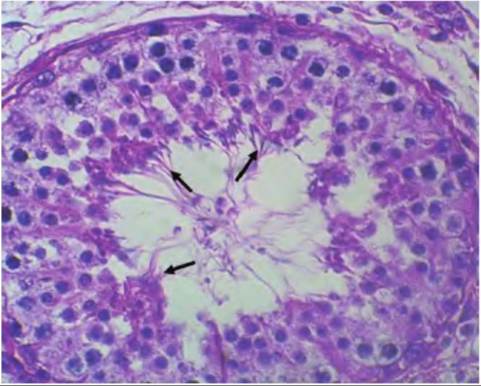
Fig. 19.20. Stage Vlll development of bull spermatogenesis.
Several clusters of mature spermatozoa (arrows) are nearing release into the lumen of the seminiferous tubule.
described as being in Stage I of spermatogenesis. The appearance is characterized by the complete absence of spermatozoa, presence of primary spermatogonia and Sertoli cells (near the basement membrane) and some round spermatids in the adluminal area, but little else.
By Stage II, some elongation of some of the spermatids begins. In Stage III, the elongating spermatids begin to clump or bunch in close association with the apical regions of the Sertoli cells. In Stage IV, secondary spermatocytes appear. Stages V-VIII are characterized by continued expansion of the epithelium until the new crop of mature spermatozoa "back out" of their docking stations with the Sertoli cells and are released so the cycle in that region of the tubule can begin once more.The peritubular cells, basement membrane, and the junctional complexes of the Sertoli cells combine to create a so-called blood-testis barrier that prevents large molecular weight materials and immune cells from entering the adluminal area of the seminiferous tubules. This is important because it prevents autoimmune reactions from impacting the developing spermatozoa as they undergo meiosis.
Sperm storage, maturation, and delivery
Once spermatozoa are shed from into the luminal space of the seminiferous tubules, the efferent ducts converge into the rete tubules and then a single epi- didymal duct. Here the cells and fluids (rete fluid) combine. The head of the epididymis is the initial segment. The head and body of the epididymis provide an environment, governed at least in part by the effects of testosterone, for maturation of the spermatozoa. Like the seminiferous tubules, the epididymis is also highly convoluted. In many species if extended it would reach 60 meters. It is surrounded by smooth muscle so that rhythmic contractions progressive propel the spermatozoa to the tail of the epididymis. Transit times vary but in bulls, for example, it takes 9-14 days for the trip from head through the tail. Once in the tail, smooth muscle activity is reduced unless there is sexual stimulation. Sexual stimulation induces smooth muscle contractions in the tail so that spermatozoa are propelled into the ductus deferens. However, transit through the body and head of the epididymis are not affected. This means that the tail region of the epididymis, especially the distal region, serves as a reservoir. Thus, sperm number per ejaculate is dramatically impacted by ejaculation frequency. In a practical sense this explains the need for not overstocking breeding males so that maximum fertility is maintained. In short, the breeding capacity depends primarily on the epididymal tail reserves rather than the rate of sperm production.
Given the continuous production of several billion spermatozoa daily there is also a continuous loss of spermatozoa irrespective of sexual activity. Since there is no reabsorption of spermatozoa, periodic contraction of the epididymis and ductus deferens produces a flow of cells into the pelvic urethra where they are flushed with urination. Figure 19.21 and Figure 19.22 illustrate histological features of the epididymis. Notice the accumulation of spermatozoa in the lumen of the caudal regions, presence of a distinct muscularis (contrast with the seminiferous tubules), and the
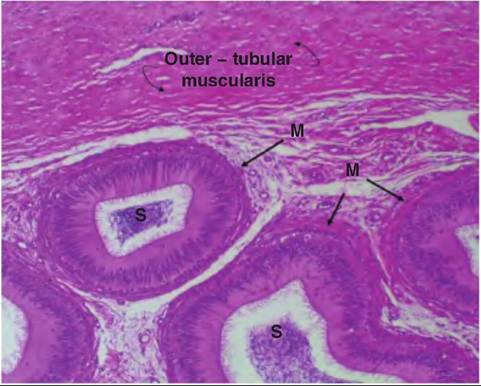
Fig. 19.21. Image of the body region of the bovine epididymis. Part of the thick muscular tunic (outer tubular muscularis) that surrounds the entire structure as well as smooth muscle (M) surrounding the highly convoluted epididymal tubular elements are shown.
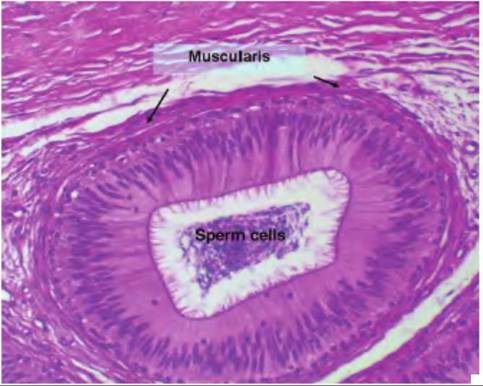
Fig. 19.22. H istological appearance of tail region of the epididymis. Note the distinct layer of smooth muscle (muscularis) surrounding the tubule, the cluster of spermatozoa in the luminal area, and the pseudostratified epithelial cells with cilia that line the internal surface.
distinct presence of the stratified, ciliated epithelium lining the internal surface of the epididymis.