Overview of skin structure
As you can appreciate, the thickness of the skin varies from region to region of the body. Consider our own bodies and the toughness of the skin on the plantar (sole) surface of our feet compared with the skin of our faces.
Now imagine how tough the skin of a horse or cow must be to withstand the environmental and physical demands. This probably explains the durability ofleather-covered furniture. However, no matter the location or thickness, skin is composed of two distinct tissue regions—epidermis and dermis. The epidermis is the outer layer and is composed of multiple layers of epithelial cells along with other specialized cells. The underlying dermis is largely composed of connective tissue and provides for the passage of blood vessels and nerves. The epidermis is also avascular. This means that nutrients must diffuse from capillaries located within the dermis to supply the epidermal cells. The tissue that lies just under the dermis is the hypodermis, often just called subcutaneous tissue. This subcutaneous tissue is not strictly part of the skin, but because the region primarily contains areolar connective tissue and adipose tissue, it serves to cushion and protect both the skin and underlying muscle and organs. Aneedle and syringe is often generically called a hypo or hypodermic syringe. Penetrating the skin and releasing the material into the space just below the dermis constitutes a hypodermic injection, hence the name.
Epidermis
The epithelium of the epidermis is a keratinized stratified squamous epithelium that consists of four cell types and four to five distinct layers (depending on location). The most common epidermal cell is the kera- tinocyte. As their name suggests, a major function of these cells is to produce keratin, a fibrous protective protein. Keratin acts to waterproof the skin and, along with secretions produced by accessory glands, protects the underlying tissues from heat, microbes, abrasion, and chemicals.
Keratinocytes are closely connected by desmosomes, which serve to anchor the cells together to physically create a more protective barrier. The keratinocytes are first produced in the cell layer closest to the underlying dermis, called the stratum basale. As the cells age, they are progressively pushed into layers closer to the surface of the body. By the time the cells reach the outermost layers, they have accumulated large amounts of keratin. The turnover of epidermal cells is rapid. The entire epidermis can be replaced every 25-50 days. In areas subjected to abrasion cell proliferation and replacement, this is even faster.Essentially, new cells are formed in the stratum basale (or stratum germinativum). This is the deepest of the epidermal layers. It is composed of one row of cuboidal to columnar-shaped epithelial cells that divide rapidly to produce new keratinocytes that are subsequently pushed toward the surface to become part of the more superficial layers. In addition, approximately 20% of the cells are melanocytes. As new cells push older cells outward, these older cells become the stratum spinosum, which typically is 8-10 cells thick. These cells contain thick bundles of intermediate filaments (Ionofilaments). In histological preparations, these cells often shrink. This causes the cells to have a prickly or spiked appearance. This explains the name of this layer, that is, the spinosum (little spine). Because the stratum germinativum and stratum spinosum are immediately adjacent to the dermis, these are the only epidermal cells that receive adequate sustenance from diffusion of nutrients from the capillaries of the underlying dermis. With further degeneration and increased keratin accumulation, the cells appear in the stratum granulosum. In this layer the keratinization process begins in earnest, and the cells begin to die. This layer is called granulosum because the accumulation of keratin granules becomes more evident in dead cells. In areas of thick skin, a layer called the stratum Iucidum can be distinguished.
Here a thin layer of cells (typically 2-3 cells in thickness) becomes translucent. With time, the dead cells, accumulated keratin, and lipids combine to create the outermost layer of skin, the stratum corneum. The relative thickness of these layers varies from region to region. Figure 5.1 provides a diagrammatic illustration of these tissue layers and Figure 5.2 and Figure 5.3 provide histological examples.There is much interest in the antimicrobial properties of molecules that accumulate in or are produced by the epidermis. For example, Schroder and Harder (1999) described an inducible, transcriptionally regulated antibiotic peptide produced by human skin. The peptide named human beta defensin-2 (hBD-2) was shown to be effective in killing gram-negative bacteria. Such findings might well find application in the dairy industry. For example, the opening of the teat end of cows and other ruminants is lined by stratified squamous epithelium, which continues into the teat meatus as the streak canal. It is well known that the
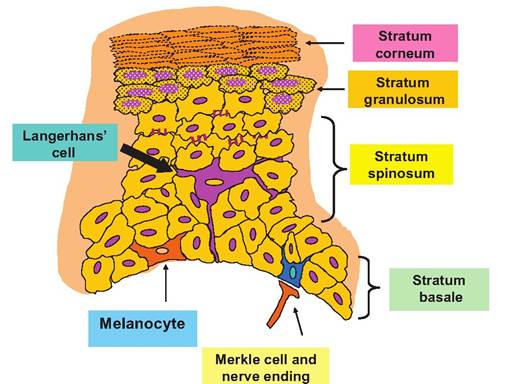
Fig. 5.1. Diagram of skin cell types and layers.
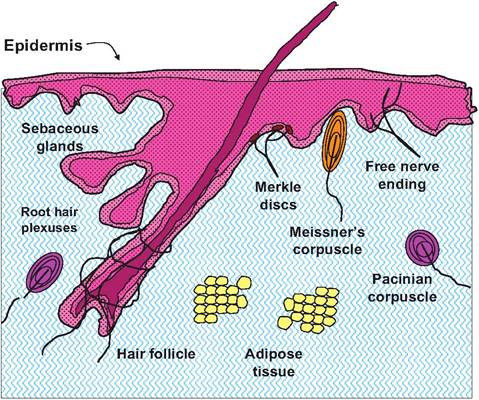
Fig. 5.3. Diagram illustrating some major features of the dermis.
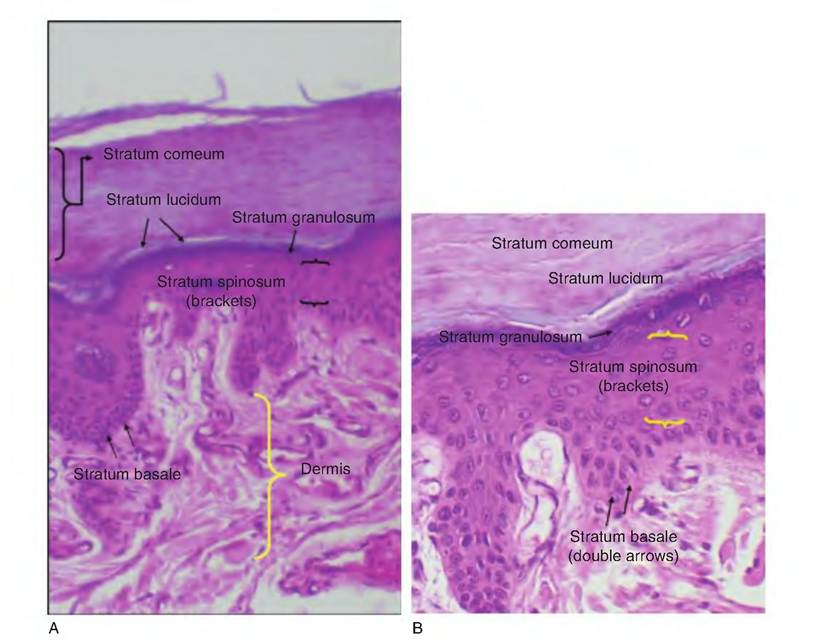
Fig. 5.2. Low-power view (A) of thick skin and cell detail (B).
keratin and other molecules serve as a barricade to seal the streak canal of the teat and protect the mammary gland from mastitis. Much of the protection involves the physical closure of the teat opening between milking episodes, but there is substantial evidence that specific components within the keratin layer can directly act as antimicrobial agents. Some of these substances are likely analogous to hBD-2, but others may be derived from mammary secretions that become trapped within the epithelium of the streak canal.
Effects might involve the direct killing of microorganisms or perhaps the prevention of colony formation.During machine milking, there are dramatic physical effects on the teat, the teat end, and the streak canal. Given that the rate of milk flow in cows is 7-8 m/ s, it is reasonable to expect that resulting shear forces might remove some of the protective keratin. It is also probable that some milk constituents are absorbed into the keratin during the time of milking or from milk droplets remaining after milking. If milking removes substantial amounts of the keratin and if renewal is delayed or changes in composition favor the formation of bacterial colonies or bacterial adherence, this could decrease the effectiveness of the streak canal as the primary defense against mastitis. In fact, experimental removal of streak canal keratin markedly increases the rate of intramammary infections. In Holsteins, keratin weight before milking was 1.6 times greater than after milking (3.1 vs. 1.9 mg per teat). Jerseys, by contrast, showed little effect of milking (3.5 vs. 3.1 mg per teat). There is a negative correlation between keratin loss at milking (r = 0.53, wet weight basis, or r = 0.65, dry weight basis) and milk production. Total lipid in the keratin is similar before and after milking. In addition, although the major aspects of the fatty acid profiles are also similar before and after milking, keratin after milking has more short-chain fatty acids. This is consistent with addition of milk-derived lipids to the keratin by contamination of milk droplets remaining in the streak canal. In the case of the Holsteins, a greater proportion of the keratin is made up of these lipids after milking (Bitman et al., 1991). Although it was once thought that the keratin regenerated rather slowly, that is, 2-4 weeks, detailed quantitative studies show that following an initial collection, the keratin regenerates at a rate of 1.5 mg (wet weight) or 0.6 mg (dry weight) per day per teat.
This suggests complete restoration of the keratin occurs within 1-2.5 days (Capuco et al., 1990). Just as in human medicine, there is considerable research interest in the identification of epidermal- keratin components that might act to protect the udder from infection. Clearly, techniques to enhance the protective function of the integumentary system would find many animal agricultural applications.In a recent paper, Fukui et al. (2012) have determined the distribution of a variety of antimicrobial products within the eccrine glands of porcine snout skin, including β-defensin 2, lysozyme, and lactoferrin. Given the propensity of pigs to root and explore, it is not hard to imagine the significance of these protective agents in secretions and surfaces of the snout.
While the keratinocytes are most plentiful, other cells also play important roles. For example, Merkel cells function as sensory receptors (touch). They orient with elements from the nervous system to create a disc-shaped sensory nerve ending called a Merkel disc. Other nerve endings and specialized receptor cells also occur in the skin, but these are located within the dermis.
A very specialized immune system cell type, Langerhans cells, develops in the bone marrow, but they migrate to the epidermis where they take up residence. They are also called epidermal dendritic cells and are essentially modified macrophages. Their name is derived from their morphology. Usually located within the stratum spinosum, they nestle between keratinocytes and send multiple projections between the cells to create an extensive network. In this way, the cells act as monitors to detect the presence of foreign debris, microorganisms, and other materials. When they are simulated, they actively process these materials and function as antigen-presenting cells to induce activity of T- and B-Iymphocytes. We have all experienced the results of immunological response in the skin after being exposed to irritants, the itch and rash.
In fact, the skin is continually exposed to an incredible variety of antigenic stimuli. Consequently, a wide array of immune responses occurs in part because of mediators secreted by keratinocytes, dendritic cells, and mast cells in the skin.Melanocytes are the final cell type of the epidermis. They are located in the lowest layer, the stratum basale, where they function to produce the pigment melanin. As melanin is synthesized, it accumulates in secretory vesicles—melanosomes—that are sequestered in elongated processes that extend from the cells. The presence of these peripheral vesicles causes the cells to have a spider-like appearance. Over time, melanin is released and taken up by surrounding keratinocytes. With increased concentrations, the melanin granules become oriented in the region of the cell that orients to the outside of the body. This pigment shields and protects the nucleus of cell from ultraviolet radiation.
Dermis
The dermis, the second major subdivision of the skin, accounts for about 80% of the total mass. It is typical of many connective tissues. There are a variety of cell types present but as you might expect, fibroblasts are common along with their products, that is, the collagen, elastin, and reticular fibers that provide essential strength and flexibility. Unlike most other connective tissues, there is also seemingly a dizzying array of specialized structures. Most of these are related to the sensory side of the nervous system. Since the integumentary is intimately associated with the external environment, various receptors provide the central nervous system with information necessary to maintain homeostasis: external temperature, pressure and touch, and presence of noxious or damaging agents. There are also other specialized epithelial structures that assist maintenance of homeostasis: sweat and sebaceous glands and hair.
The dermis consists of two layers: the papillary layer and the reticular layer. The papillary layer is the outer region closest to the epidermis. In this area, there are fingerlike projections called dermal papillae (these also give the layer its name) that penetrate into the epidermis. In some areas, for example, the palms of the hands or fingertips in humans and apes or the pads of a cat's foot, the papillae are arranged on the top of larger structures called dermal ridges. This acts to increase friction and allows for an easier grip. The particular pattern of ridges is unique to each individual and is the basis for fingerprints in humans or other primates. These projections contain capillaries, and a variety of nerve endings and receptors. Three broad groupings of receptors include (1) exterocep- tors, (2) Interoceptors (sometimes called visceroceptors), and (3) proprioceptors. Exteroceptors are concerned with stimuli that arise from the outside. Most extero- ceptors are located on or near the body surface. These are the focus of our study of the integumentary system. Interoceptors react to stimuli from within the body, for example, chemical signals, temperature, or gut motility. Proprioceptors also reflect internal responses but are specifically involved in the relay of information concerned with muscle, tendon, or ligament movement or stretch. In other words, they monitor the musculoskeletal organs. The latter two classes of receptors will be discussed in subsequent sections. Our focus now is on exteroceptors of the skin.
These receptors can also be classified based on their structural complexity. Free nerve endings, for example, are structurally simple, especially when compared with receptors associated with the special senses (vision, hearing, olfaction, or taste). Even the relatively simple receptors of the integumentary can be divided into unencapsulated (free nerve endings) and encapsulated groupings. Specialized Meissner's or Pacinian corpuscles are examples of encapsulated receptors. Pacinian corpuscles have a structure similar to the layers of an onion; pressure induces ion changes that are translated into graded potentials in the associated nerve fibers (Fig. 5.5). These impulses are interpreted as touch or pressure by neurons in the cerebral cortex. Table 5.1 summarizes the types and classes of sensory receptors within the skin.
The reticular layer is the thicker and deeper layer of the dermis. It is composed of dense irregular connective tissue and contains thick bundles of interlacing collagen fibers and some coarse elastic fibers. However, elastin fibers are typically only visible after special staining. These fibers run in several directions, which increases strength, but most are oriented parallel to the skin exterior. The fibers provide much of the strength and resistance to stretch in skin and the long- wearing attributes of leather. The reticular layer is also abundantly supplied with blood vessels and nerves. These elements of the dermis are illustrated in Figure 5.3. Figure 5.4 and Figure 5.5 show histological examples of some of the other structures that occur within the dermis.
Specialized structures
Sweat glands
The dermis contains a variety of glandular structures. Sweat glands in primates are plentiful and widely distributed. The most common type is the eccrine
Table 5.1. Integumentary sensory receptors classified by structure and function.
| Structural Class | Functional Activities | Location |
| Unencapsulated Free nerve endings | Nociceptors (pain); thermoreceptors; mechanoreceptors (pressure) | Most tissues |
| Modified nerve endings (Merkel discs) | Mechanoreceptors (light pressure) | Stratum basale |
| Root hair plexuses | Mechanoreceptors (hair movement) | In and around hair follicles |
| Encapsulated Meissner's corpuscles | Mechanoreceptors (light pressure, discriminative touch, vibration) | Dermal papillae, esp. face, fingertips |
| Kraus's end bulbs | Mechanoreceptors (modified from Meissner's corpuscles) | Connective tissue of mucosae |
| Pacinian corpuscles | Mechanoreceptors (deep pressure, stretch, rapid adaptation) | Widespread in skin |
| Ruffini's corpuscles | Mechanoreceptors (deep pressure, stretch, slow adaptation) | Deep dermis, joint capsules |
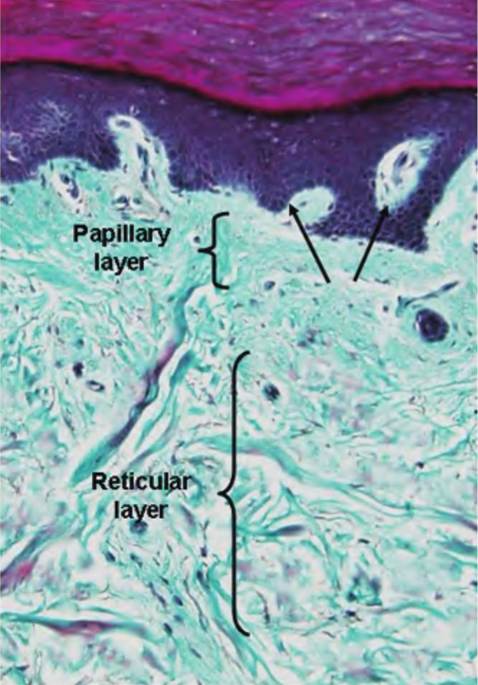
Fig. 5.4. Example of thick skin. In this preparation the dermis is stained a pale turquoise and stands in sharp contrast to the epidermis. Arrows indicate dermal papillae.
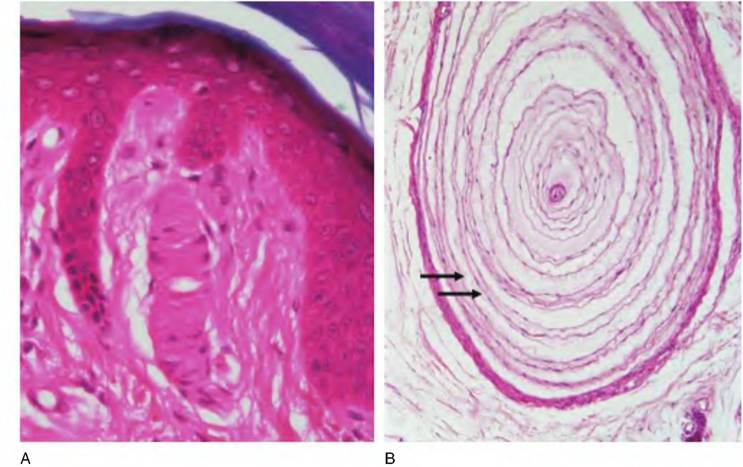
Fig. 5.5. Structure of Meissner's corpuscle (A) and Pacinian corpuscle (B). Both of these sensory receptors are located in the dermis and are responsive to pressure and touch. Arrows indicate multiple layers of a Pacinian corpuscle.
gland. These simple coiled glands open onto the surface in pores. They produce a hypotonic watery secretion that derives from interstitial fluids. It is mostly water with some dissolved salts, lactic acid, and traces of other waste products. The rate of secretion is controlled by the activity of the sympathetic nervous system. In humans, a typical response occurs with overheating. Sweating induced in this way begins on the forehead and progresses downward. Emotionally induced sweating—due to fright, embarrassment, or nervousness—begins on the palms, soles, and armpits, and spreads to other areas. Of course, the primary function of sweat is to cool the body as a result of evaporation. A second type of sweat gland, apocrine glands, makes up a small proportion of the total. These glands are larger than eccrine glands, and their ducts open onto hair follicles. They are primarily confined to the axillary and anogenital areas of the primate body. The secretions in addition to watery sweat also contain fatty acids and some proteins. These glands are affected by sex steroids; that is, activity begins with the onset of puberty. For this reason, apocrine glands are believed to be analogous to the scent glands of other animals. Eccrine sweat glands are sparse among domestic animals. For example, in dogs and cats they are located only on the footpad. This limited distribution means that these glands have virtually no effect on heat loss, but they do act to moisten the surface and improve traction. We have all noticed the panting dog on a hot day or after exercise. Panting is an effective cooling mechanism because it moves greater amounts of air over moist surfaces. This extra water-saturated air is exhaled, and in the process, body temperature decreases.
Although eccrine sweat glands are lacking, horses, cattle, sheep, swine, dogs, and cats have numerous apocrine glands. In the dog, for example, the proteinaceous, whitish secretions from the apocrine glands mix with the oily secretions of the sebaceous glands to form an emulsion-like coating on the skin. The characteristic dog, horse, or cow odor is primarily a result of bacterial action on these accumulated secretions. These secretions also impact heat loss, but this is likely most effective in horses, followed by cattle, sheep, dogs, cats, and swine. Regardless of the evaporative effect of heat lost from sweat, the skin of these animals is nonetheless important in temperature regulation. This is because simply changing the rate of blood flow through capillaries in the skin alters the volume of warm blood near the surface of the body, thereby affecting thermoregulation.
Sebaceous glands
Sebaceous or oil glands also occur in mammals. These are simple branched areolar glands that release their products (holocrine mechanism) onto the hair follicles. The secretion is called sebum, which is a mixture of cellular lipids and other cell components. Sebum is a natural skin cream and hair protector. It helps keep hair from becoming brittle, prevents excessive evaporation of water from the skin, keeps the skin soft, and contains a bactericidal agent that inhibits the growth of certain bacteria. This is closely related to lanolin that secretes onto wool fibers in sheep.
Other skin glands
Many animals have glands associated with the anal region. These glands are usually divided into three classes depending on specific location and orientation: (1) anal glands, (2) glands of the anal sac, and (3) perianal glands. Anal glands are found in dogs, cats, and pigs. They are modified tubuloalveolar sweat glands located in the submucosa of the anal canal and the opening of the anus. Carnivores secrete a Iipid-Iike material, but pigs secrete a more mucus-like material from these glands. Clusters of lymphatic tissue, similar to the Peyer's patches in the intestine, often accompany the glands.
Anal sac glands, sometimes called perianal sinuses, occur in pairs and are essentially invaginations or diverticula of the anal surface. They are Iocatedbetween the outer and inner anal sphincters. Within each pocket or sac, glands embedded in the wall have openings that release the contents into the space of the sac. The anal sac is present in carnivores and rodents. In dogs, the glands are arranged as compound tubular structures and exhibit an apocrine mode of secretion. In cats, the glands are similar, but apocrine and holocrine secretion occurs. Products from the glands open into secretory ducts in the neck of the anal sac. The excretory secretions of the anal sac glands, sloughed cells, and fecal material can block the openings of these anal sacs. When the sacs become blocked, they may have to be manually expressed—not a pleasant experience.
Circumanal or perianal glands are anomalies in the sense that they often appear as masses of epithelial cells within the submucosa that appear not to have functional ducts that lead to the surface. They are sometimes oriented adjacent to sebaceous glands, which suggests they may be related, but this is far from certain. These nondescript masses of cells are frequently described as nonsecretory glands. Unfortunately, the solid masses of cells are believed to be especially prone to neoplasia.
The mammary glands are also skin glands but will be considered in greater detail in a subsequent chapter. Some other specialized glands include the infraorbital glands of sheep, the submental organ of the cat, and the scent or horn glands of goats. For example, the submental glands of the cat are located within the Intermandibular space under the jaw. The "organ" is essentially a cluster of sebaceous glands. It is not uncommon to note domestic cats marking their territory by rubbing their chins. In goats, this activity is a bit more apparent. The scent glands in the goat are located along the caudal to medial aspect of the base
Box 5.1 Skin derivatives
Every biology student quickly learns that the presence of functioning mammary glands is a hallmark of mammals. As outlined in his recent review, Oftedal (2012) provides support for the idea that primitive lactation began as a reproductive advantage in synapsids (the direct ancestors of mammals) likely during the Pennsylvania period. The capacity of "modern" mammary glands to provide secretions with both vital nutrients (proteins, fats, and carbohydrates) and health benefits (immunoglobulins, lysozyme, etc.) likely evolved from apocrine- like glands that were associated with hair follicles. It is believed that secretions from these glands provided moisture and antimicrobial agents for the parchment-like shelled eggs laid by these ancestors. Fossil evidence suggests that some therapsids and mammaliaformes present during the Triassic period produced milk-like secretions. Evolutionary pressure likely promoted the incorporation of molecules such as lysozyme or iron-binding lactoferrin into these secretions. Certainly, the capacity to prevent desiccation of the eggs and protection from microbial attack would have been highly beneficial. Thus, the significance of the skin and skin-derived molecules and skin-derived glands remain physiologically critical in humans and animals.
of the horns. Rubbing this area leaves sebaceous secretions that are apparent because they contain caproic acid. This is the short-chain volatile fatty acid that is responsible for the distinct odor of male goats. The uropygial gland of birds, also called the oil or preen gland, is the only skin gland of birds. It is composed of a series of closely aligned sebaceous adenomeres (secretory units) that empty into a common space or sinus that ultimately empties onto a common papilla. The papilla has associated smooth muscle fibers that surround the duct opening. The opening is located above the last sacral vertebra (Box 5.1).
Hair
Functions associated with hair include insulation, protection, and sensory reception. Hair or fiber production in domestic animals is directly related to the number and size of the follicles in the skin. As you might guess, various aspects of hair growth and development have been extensively studied in sheep and goats. Regardless of whether the animal in question is used for fiber production or not, unlike that of humans, the hair of animals is especially important physiologically. We will begin by first considering some of the basics of hair structure and properties. Hairs or pili are
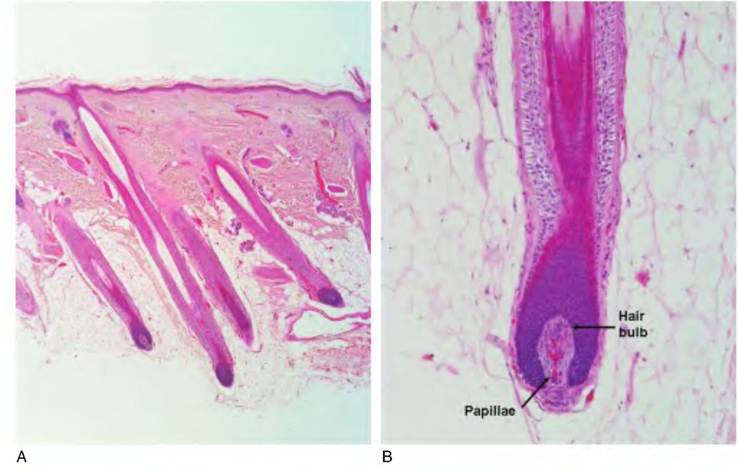
Fig. 5.6. Histology of hair follicles. Histological section of several hair follicles at low magnification (A) and of a single follicle at higher magnification (B) is shown. At lower power it is evident that the hair bulbs penetrate deep into the dermis and hypodermis.
generated by hair follicles and are flexible strands that mostly consist of layers of dead keratinized cells. However, the keratin in hair is harder and more durable than the softer keratin of the epidermal cells. The parts are simple: (1) the shaft projects from the skin; (2) the root is embedded in the skin. In humans, if the cross section of the shaft is ribbonlike, the hair is kinky. If the shaft cross section is oval, the hair is wavy. If the shaft cross section is round, the hair is straight and relatively coarse. These basic relationships also apply to animal hair.
At a more detailed level, the hair shaft can be divided into three regions: (1) outer cuticle, (2) inner cortex, and (3) a central medulla. The outer cuticle is a layer of cornified epithelial cell "husks" that closely interlink with the cuticle cells of the root of the hair. The cortex makes up the majority of the hair shaft and is composed of several layers of flattened cornified cells that have accumulated "hard" keratin. Pigments may also appear in these cells along with air spaces. The medulla contains cells that are more cuboidal. These layers are especially clear in cross-sectioned hair shafts. The hair terminates in the root or hair bulb. Essentially, the hair resides in an invagination of the surface of the epithelium that extends to the dermis. This is important because it allows ready transfer of nutrients and waste products from the hair bulb to the interstitial fluid of the dermis. Growth occurs when cells in the apex of the root bulb give rise to new medullary cells. Laterally positioned cells give rise to the cells of the inner cortex and outer cuticle, respectively. The growth of the hair then is analogous to the growth of the epidermis generally. Cells from lower depths progressively displace those above. See Figure 5.6 for examples of hair and hair follicle structure.
In many situations there is a sheath of smooth muscle, the arrector pili, that attaches to the connective tissue that surrounds the hair follicle and a portion of the hair shaft that is underneath the surface of the skin. The contraction of this smooth muscle causes the hair to stand on end and is likely associated with increased secretion of surrounding sebaceous glands. This is the basis of a hair-raising experience or the familiar Halloween cat with its arched back and raised hair.
In horses and cattle, the hairs are evenly distributed across the body, but in other species (dog, cat, pig), the hairs are oriented into groups called hair beds. In the dog, each hair bed has a group of follicles that consist of one larger major hair (guard or principle hair) typically about 150 μm in diameter. A cluster of auxiliary hairs that are typically shorter and only about 75 μm in diameter surrounds the guard hairs. These hairs often exit the skin in the same opening as the guard hairs. In addition, the auxiliary hairs do not have a medulla. Clustering of hairs in this manner can be extreme. For example, the chinchilla, noted for its soft, dense pelt, can have clusters that have 50-75 auxiliary hairs each. In this case, the guard hair is also very fine and only slightly larger than the auxiliary hairs, that is, 15 versus 11 μm in diameter.
Other specialized hairs include tactile or sinus hairs. These are familiar as cat's whiskers, for example. These hairs are usually longer and larger than normal but share similar structures to normal guard hairs. The roots of these hairs are highly innervated by free nerve endings and Merkel's discs.
The hair growth cycle is divided into three phases: (1) anagen, (2) catagen, and (3) telogen. Hairs are generated by the proliferation of cells within the hair bulb during anagen. The continuous addition of new cells to the shaft or the hair produces elongation. Termination of growth occurs when mitotic activity of the basal of germinal cells of the bulb decreases. Catagen is a transition phase. It is characterized by gradual transition of the bulb cells. The cells progressively convert
into a solid, keratinized mass, and the more distal region of the follicle thins. The bulb is forced toward the surface and the papilla is lost. In this condition, the hair is referred to as a club hair. After a time, a secondary germ structure develops deep under the club hair. The formation of the new germinal center marks the beginning of telogen, which can last for weeks or even months. During early anagen, the new hair bulb progressively elongates and the shaft of the new hair displaces the older club hair. Figure 5.7 provides a diagrammatic illustration of the cycle of hair growth.
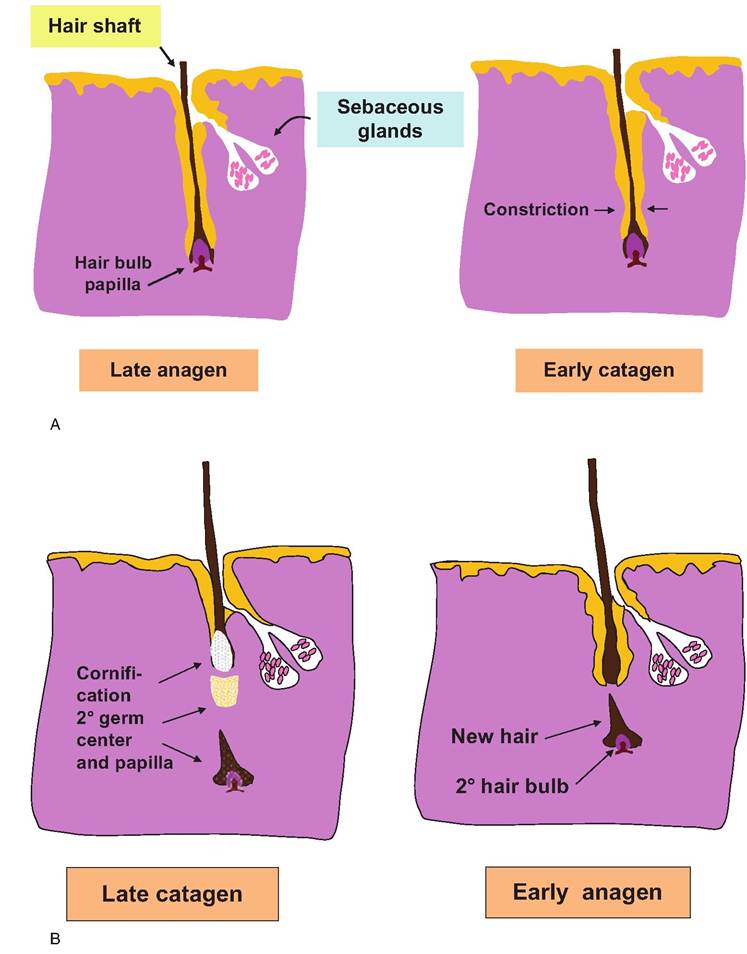
Fig. 5.7. Diagram of hair cycle events. Growth begins to slow in late anagen, followed by constriction of the follicle in early catagen. By late catagen, the hair is increasingly cornified, producing a club hair that is progressively extruded. A 2o germ center and papilla appear some time after. By early anagen, a new hair bulb and growing hair shaft begin to develop. The new hair often follows the same path as the old hair follicle.
The follicle population determines the quantity and quality of wool production. As a general rule, a high number of hair follicles leads to production of fibers with a lower diameter but the opposite with low follicle density, that is, fibers that are thicker and more coarse Hocking Edwards et al., (1996). It is also apparent that hair growth is cyclic but that this basic rhythmic pattern can be modified by external cues. This is evident in the seasonal pelage cycles noted in many mammals. Recent studies have focused on the role of the pituitary hormone prolactin in wool growth, for example. The secretion of prolactin is well known to be altered by changes in photoperiod, specifically decreased secretion during short days, and typically in cooler temperatures in regions well above or below the equator, with increased secretion during long days and warmer temperatures. Reduced photoperiod and correspondingly reduced prolactin concentrations in the blood are associated with the stimulation of hair growth and generation of the winter pelage. Nixon et al. (2002) reported changes in expression of prolactin receptor within the dermal papilla and the outer root sheath of wool follicles of sheep. Moreover, changes in receptor expression were altered in sheep subjected to photoperiod-induced changes in circulating prolactin. This suggests that changes in expression of prolactin receptor within specific regions of the follicle are involved in seasonal changes in hair growth.