Biochemical properties of skin
In this section we will consider physiological attributes of the integumentary system. Included here are aspects related to vitamin D synthesis and homeostasis. We also discuss factors that control coat and skin color.
Skin and coat color
In humans, three primary pigments give color to the skin. Melanin is especially important and can produce various shades of yellow, brown, or black. The rate of melanin synthesis is largely genetically determined. For example, if you have very fair skin, melanin production is low, and it is a very light shade. If you have very dark skin, on the other hand, melanin production is higher, and the pigment is inherently darker. It is believed that the average density of melanocytes is relatively constant between people, but rates of cellular activity vary. The yellow-orange pigment carotene also impacts skin color. It tends to collect in the stratum corneum and in the adipose tissue of the hypodermis. Hemoglobin, the oxygen-carrying protein of red blood cells, also impacts skin color. To illustrate, a blush, especially in a fair-skinned person, is a result of a quick flush of oxygenated blood through the capillaries of the dermis of the cheek. Clinically, the appearance of a bluish cast to the skin or nail beds is an indication of inadequate oxygenation of blood, perhaps as a consequence of anemia or some other problem, such as a respiratory or cardiac problem. Other items related to skin color include (1) jaundice, caused by deposition of bile pigments in certain liver diseases, (2) hematoma, the blue-black color produced by bruising when blood vessels in the skin rupture, and (3) erythema, the term for the reddish cast produced by blushing, fever, or strenuous exercise.
What about skin or hair color in animals? In reality, there are two subtypes of melanin: eumelanin (brownblack) and pheomelanin (yellow-red).
The making and processing of melanin, melanogenesis, is complex because of interactions of multiple cell types in the skin. In mice, for example, nearly 100 genes are known to affect coat color. However, these genes are classified into two primary groups: (1) those that act on the melanocyte and (2) those that directly impact the biochemistry of pigmentation. The melanocytes residing in the stratum basale and within hair bulbs synthesize and package melanin into secretory vesicles called melanosomes. The melanosomes are then distributed to surrounding cells. It is easy to imagine sources of variation in these steps and how this might affect skin or hair color. For example, which type of melanin is made, how much is made, where it is distributed, how quickly it is degraded, and so forth. Tyrosinase is the essential regulatory enzyme in melanin synthesis. With high rates of enzyme activity, the formation of eumelanin is enhanced; lower rates favor formation of pheomelanin. In addition to synthesis variation, melanocortin receptors are critical in initiation of melanin synthesis. Hormone control of pigmentation is evident in many situations, for example, changes in fur color in Arctic mammals and increased skin pigmentation, which is a symptom of primary adrenal gland dysfunction. Melanocortin refers to a large family of structurally related hormones derived from the precursor protein pro-opiomelanocortin (POMC). This protein is abundant in the intermediate lobe of the pituitary gland. Depending on processing, at least four melanocortin peptides can be generated. These are proteins that can interact with the melanocortin 1 receptor (MClR). These include adrenocorticotrophic hormone (ACTH), as well as oc-, β-, and γ-melanocyte- Stimulating hormone (MSH). Excess production of ACTH explains the excess skin pigmentation that occurs with adrenal insufficiency disease, since ACTH readily binds to the MCl-R receptor on the melanocytes. MCl-R is a G-protein-linked receptor (see endocrine chapter) with seven transmembrane spanning domains. However, variants of the receptor (MC 2-, 3-, or 4-R) are expressed in other tissues. This suggests that| Table 5.2. Mutations, gene products, and phenotypes related to pigmentation genes in mice. | ||
| Mutation | Gene Product | Phenotype |
| Albino | Tyrosinase required for melanin synthesis | Absence of pigmentation |
| Agouti | Secreted ASP, agonist of MC1-R | Loss of function = black hair Gain of function = yellow hair |
| Brown | Tyrosinase-related protein 1 | Loss of function = brown instead of black hair |
| Lethal spotting | Endothelin 3 (peptide related to angiogenesis) | Piebald spotting, deafness, death |
| Piebald spotting | Endothelin receptor type β | Same as for lethal spotting |
| Pink-eyed dilution | Integral membrane protein in the melanosomes | Loss of function = pink eyes, yellow-gray hair |
| Recessive yellow | Melanocortin receptor | Loss of function = yellow hair Gain of function = black hair |
| Slaty | Tyrosinase-related protein 2 | Partial loss of function = dilution of black hair |
| Steel | Mast cell growth factor | White hair, black eyes, sterility |
| White spotting | Receptor tyrosine kinase (encoded by the proto-oncogene kit) | Partial loss of function = piebald spotting |
the melanocortin ligands have a variety of physiological effects in addition to pigmentation of skin or hair.
Mouse coat color genes were some of the first mutations discovered that have been related to regulation of coat color. Resource animals for many of the classic mammalian genetic studies have their foundations in the very large numbers of coat color variations (brown, silver, and yellow) that were described by fanciers of unusual mice in Europe and Asia in the 18th and 19th centuries. During this time, animals with unusual or striking variation in their pelage were prized and consequently saved as breeding stock. These fancy mice provided many of the initial resources to generate strains of mice that are used extensively in research today. Of course, understanding of molecular basis for variations in these traits has mushroomed in recent years. Table 5.2 summarizes some selected mutations and corresponding phenotypes related to pigmentation in mice.
It is now recognized that most of these mouse genes have counterparts in other animals. For example, black horses are homozygous for a deletion in the agouti locus. The mutation producing the chestnut allele is a single base substitution on the MCl-R gene (Rieder et al., 2001). In domestic pigs, the predominant white phenotype is linked with two mutations in the KIT proto-oncogene, which encodes for mast (stem) cell growth factor receptor. The many millions of white pigs around the world are assumed to be heterozygous or homozygous for these two mutations (Marklund et al., 1999). In the bovine, three alleles of the MSH receptor gene (localized to chromosome 18) have been reported. A point mutation in the dominant allele ED results in black coat color. However, a frameshift mutation, which causes a synthesis of a shortened receptor protein in homozygous e/e animals, leads to a red coat color. The wild-type allele E+ allows production of a variety of colors. This likely reflects the great variety in regulation of the normal receptor (Klungland et al., 1995).
In mice, the switch between production of eumela- nin and pheomelanin production is tied not only to the binding of the melanocortin ligands to MCl-R, but also to signaling from the agouti signaling protein (ASP).
ASP is a soluble paracrine protein made by dermal papilla cells within the hair follicles. Binding of ASP to MCl-R prevents normal binding of OC-MSH and thereby promotes production of pheomelanin rather than eumelanin. In mice with the agouti phenotype, the promoter of the ASP gene is transiently activated during the mid-phase of the hair growth cycle. This causes a band of yellow pigment (accumulation of pheomelanin) in hair that is otherwise black (eumelanin accumulation). Strains of mice with entirely yellow hair coats have mutations that involve this ASP gene. For example, in one case, a prevailing mutation in the agouti locus (Ay∕ A) results in excessive production of ASP throughout the body. This causes the complete yellow hair coat as well as obesity. The obesity results from ASP antagonism of the MC4-R receptor in the hypothalamus. In another strain, the complete yellow coat is produced in a recessive allele (e/e), in the gene for MCl-R. In these animals the receptor protein is synthesized with extra amino acids (extension locus), which causes a failure of signaling. This leads to the exclusive production of pheomelanin and the total yellow hair color. Interestingly, yellow mouse hair is thought to be the murine equivalent of red hair in humans (Schaffer and Bolognia, 2001).As emphasized in recent reviews (Schneider et al., 2009; Lee and Tumbar, 2012), molecular understanding of hair follicle physiology has depended on study of various mouse mutants (either naturally occurring or genetically engineered) that express various abnormalities in hair structure, growth patterns, and/or color. Moreover, a detailed study has also provided general insights related to topics such as organogenesis, regeneration, morphogenesis, stem cell biology, cell proliferation, and cell migration, as well as apoptosis. As outlined by Lee and Tumbar (2012), key cell-signaling
Box 5.2 Hair follicles to the rescue
While we have indicated the significance of the skin in protection against disease, as described by Heath and Mueller (2012), it may be that hair follicles are also important players in the recruitment of anti- gen-sensing dendritic (Langerhans) cells to the epidermis of the skin.
Nagao et al. (2012) have shown that recruitment of bone marrow monocytes to become dendritic cells depends on hair follicles. In their study, they we able to induce specific depletion of Langerhans cells from the skin and then monitor reestablishment. To their surprise, in addition to blood-derived monocytes, precursor cells appeared to reside in the hair follicle. Regardless of the source (blood or follicles), cells that become the newly derived dendritic cells gained access to the epidermis via the Interfollicular epidermis of the hair follicles. They used histology and multiphoton microscopy to identify the cells and track their migration. In support of the role of the hair follicles, the skin of mutant mice that do not produce hair follicles has little capacity to repopulate the skin with dendritic cells when they are depleted. Repopulation of areas without hair follicles, that is, the foot pad, depends on adjacent regions with hairy skin. Finally, gentle abrasion of the skin (removal of adhesive tape) induces a rapid accumulation of neutrophils and dendritic cells around the follicles. These results suggest important roles for hair follicles in regulation of migration between the dermis and epidermis and the role of chemokines produced in the follicles as regulators of trafficking of cells and of the immune responses of the integumentary system.families involved in hair follicle development and physiology include Wnt-related, BMP∕TGFβ-related, Shh-related, and EGF/FGF-related, as well as multiple transcription factors. Also, many questions remain. How are signals from the dermis, adipocytes, or blood vessels controlled? How can these networks of signaling pathways be regulated to treat diseases or perhaps alter wool and hair production commercially? (See Box 5.2).
Vitamin D metabolism
Vitamin D has three major effects: (1) promote absorption of calcium from the intestine, (2) activate resorption of calcium from bones, and (3) increase excretion of phosphate via the kidneys. In conjunction with increased concentrations of parathyroid hormone, there are increased amounts of biologically active vitamin D to increase blood calcium. Thus, overall health and
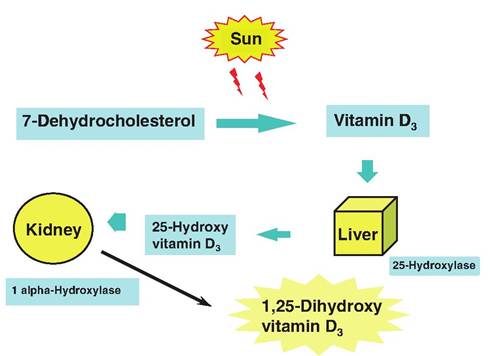
Fig. 5.8. Metabolism of vitamin D.
homeostasis require this vitamin. This story begins in the skin. Vitamin D2 is a plant product generated by ultraviolet irradiation of ergosterol. This precursor can appear in the diet. However, vitamin D3 or cholecal- ciferol is also synthesized in the skin. This is induced by the UV Irradiationof 7-dehydrocholesterol. However, before the vitamin D3 is fully active, it must be modified. The first step occurs in the liver with the conversion of vitamin D3 into 25-hydroxycholecalciferol by the addition of a hydroxyl group by the action of the enzyme 25-hydroxylase. This material again enters the bloodstream and is taken up by the kidney cells. A second hydroxyl group is added to create the most potent from of vitamin, 1,25-dihydroxycholecalciferol. This requires the actions of the 1 «-hydroxylase. This form of the molecule acts on the intestinal epithelial cells to stimulate the synthesis of calcium transporter molecules that increase the absorption of calcium.
Deficiencies of vitamin D can lead to malfunctions in calcium homeostasis so that bone growth and development is impaired in growing animals. This produces rickets, a syndrome characterized by bowlegs or knock-knees. In human medicine, as the importance of vitamin D was realized, this resulted in the practice of fortifying milk with vitamin D to help ensure that children received adequate amounts of the vitamin to minimize the appearance of rickets. Interconversions of vitamin D are illustrated in Figure 5.8 (Box 5.3).