Reproductive technologies
A variety of techniques often called assisted reproductive technologies (ART), for example, in vitro fertilization (IVF), superovulation, ET, and freezing, are popularized because they are used to assist couples incapable of conceiving naturally.
However, these tools have their origins in reproductive physiology of laboratory and farm animals. For example, scientists worked for many years to perfect IVF before it was recognized that sperm cells have to undergo capacita- tion in the female reproductive tract before fertilization. This ultimately led to methods to induce Capacitation, and therefore routinely successful IVF Advances in one aspect of reproduction thus allow progress in other areas. For example, ET would not be practical without endocrine tools to induce superovulation and methods to routinely collect, fertilize, and freeze eggs. Cloning and development of transgenic animals depended on development of techniques to visualize, micro-manipulate, and culture eggs and embryos. Several of these key techniques are outlined in subsequent sections (Bearden et al. 2004).Artificial insemination
The artificial insemination (AI) industry is an excellent example of a practical link between scientific understanding, rapid adoption of practical procedures, and creation of an entirely new agricultural industry. The foundations to understanding semen preservation and storage did not occur overnight. However, it was clear, especially to dairy producers, that techniques to select and preserve semen from genetically superior bulls would rapidly increase genetic progress. Specifically, a highly selected sire could potentially be mated with thousands of females. The first commercial AI using fresh semen dates back to 1937, but this was built on a long history of discoveries.
For example, in 1677, Leuwenhoek was the first to see spermatozoa with his newly created microscope.
The Italian scientist Spallanzani discovered in 1780 that a dog could be impregnated with the cellular portion of semen and that semen could be cooled and used at a later time. The Russian professor Ivanov developed techniques for collection of semen and insemination of cattle, horses, sheep, and pigs by the early 1930s. A Danish scientist, Sorenson, who had connections with Ivanov, established the first AI cooperative for dairy producers in 1933.The extension dairy specialist E.J. Perry from Rutgers University was visiting Denmark at this time and returned to establish the first AI cooperative in the United States in 1937. This exploded, with the creation of seven AI cooperatives by 1939. This pace of development continued as more AI organizations appeared and the number of cows inseminated increased dramatically. To control costs, many small cooperatives ultimately consolidated.
It is estimated that 60% of U.S. dairy cows are artificially inseminated. In contrast, more than 90% of dairy cows in Europe are bred using Al. The two largest AI organizations are privately owned companies with national and worldwide semen distribution efforts. The largest of these, American Breeders Service Global, sells approximately 3.5 million breeding services per year. The second largest is Select Sires.
Although methods for collecting semen and insemination have been available for many years, it is likely that the commercial industry would not have developed as rapidly without several critical research discoveries: (1) the development of semen extenders that would protect sperm cells against temperature shock and thereby allow cold storage, (2) the realization that bull semen could be extended (diluted) to breed large numbers of cows from each ejaculate, and (3) the discovery of methods for frozen storage of bull spermatozoa. In 1939, Drs. Phillips and Lardy at the University of Wisconsin discovered that egg yolk would protect sperm cells from temperature shock upon cooling.
The protection was due to phospholipids and lipoproteins in the egg yolk. Extenders combining egg yolk with phosphate, citrate, and bicarbonate buffers were soon developed and they form the basis for extenders in use today. Heated milk was also found to be a satisfactory semen extender and to provide temperature shock protection.Spermatozoa were some of the first cells frozen by English scientists in 1949. Dr. Polge and coworkers discovered that glycerol in the extender media would protect fowl and bull spermatozoa from damage during freezing. Early freezing and storage was accomplished with dry ice and alcohol at a temperature of -79oC. Afterward, liquid nitrogen became the coolant of choice because its -196oC temperature provided longer and safer storage conditions. The first U.S. calf (named Frosty) from frozen semen was born in 1953.
Dairy producers use AI in their herds because of the following advantages: (1) ready access to genetically superior sires, (2) ability to maintain a herd closed to new animals for added disease protection, (3) a dangerous bull need not be kept on the farm, and (4) organized breeding management and record services are provided. In swine operations, AI is often used to decrease the cost of maintaining males at the production unit. In turkeys, AI is used because mating cannot occur naturally because of development of very largebreasted birds.
The techniques are generally available for use of AI in other species, but use is limited for economic reasons or in horses because the registry of offspring is restricted. Changes in industry structure, however, often drive changes in the use of Al. For example, in the swine industry, the growth of large farm units in the 1990s led to the adaptation of AI to reduce expenses of maintaining males. In the horse industry, a change in the Quarter Horse Association regulations to allow the registration of foals from AI with cooled and shipped semen has increased the use of Al.
In vitro fertilization (IVF)
The relevance of in vitro fertilization (IVF) as a research tool has been recognized for many years. The first report of offspring (rabbits) produced this way was in 1959. The first "test-tube" human infant was born in 1978. In cattle, successful fertilization occurs more readily with oocytes that are ovulated compared with those collected from ovarian follicles. Regardless, this in part explains the importance of collecting eggs from animals that are superovulated (hormonally treated so that a greater than normal number of eggs are produced and ovulated) for both ETs and cloning (i.e., genetically superior donors), as well as donors of eggs to be used in transgenic trials.
Methods to improve techniques for oocyte collection and culture conditions to produce a greater percentage of successful oocytes collected from ovaries are under active development. These in vitro maturation (IVM) schemes are efforts closely tied with ultrasound-guided follicular aspiration, also called Iransvaginal ovum recovery. In this method, an aspiration needle inserted through a guide within a trans- vaginal ultrasound probe is passed through the vaginal wall and into each follicle while applying a vacuum. Typically, six to eight oocytes can be collected per ovary following superovulation. In theory, regardless of superovulation, it is possible to recover 10-20 oocytes every 10-14 days. Current IVM and IVF would be expected to produce three to five embryos suitable for implantation.
The advantage of this system is the ability to collect oocytes from cows of known genetic merit, at least twice weekly for extended periods of time, as source material for IVF and microinjection of DNA (see below). This procedure has been shown to yield about 10% greater development of microinjected zygotes to the blastocyst stage and a greater percentage of transferred blastocysts developing to term compared with use of slaughterhouse-derived oocytes (Krisher et al., 1995).
Embryo transfer (ET)
Although embryo transfer (ET) has been in the popular press recently because of implications to human reproduction, the basic techniques have been known for many years. The first recorded successful case in 1890 by the physiologist Walter Heap was done with rabbits. By the 1930s, transfers with sheep and goats were reported and in the 1950s, the first successful ETs in cattle occurred in Cambridge, England. The first successful ET in cattle in the United States was in 1951. In the United States, commercial ET work began in the 1970s. The first Holstein ET calf was registered in 1974. The popularity of ET in the dairy industry increased dramatically with development of nonsur- gical methods for recovery of embryos and subsequent implantation of either freshly collected or frozen embryos. The first successful nonsurgical embryo collection was reported in 1964 and the first North American calf from a frozen embryo was in 1977. Further development of the ultrasound Iransvaginal collection method combined with improvements in IVM will likely further increase use of ET to expand populations of genetically superior cattle or to allow for cloning of transgenic animals. At the current time, approximately 80% of young AI sires are produced by ET; approximately 12,000 ET heifers are registered each year.
Finally, it is also possible using PCR techniques designed to amplify male-specific sequences on the Y chromosome to determine the sex of embryos. Briefly, only a small sampling of cells from the blastocyst (perhaps only one cell) can be utilized. The detection of these sequences in the amplified DNA from these cells would indicate a male and the absence a female. Biopsied cells are quickly reproduced. In France, where this technology has been rapidly adopted, many dairy producers will no longer transfer embryos without having them sexed.
Sexed semen
Since the initiation of Al, countless numbers of techniques have been investigated with the goal of separating spermatozoa into X- and Y-bearing cells as a means to control sex of offspring.
Most of these methods were only marginally better than chance and suffered from poor repeatability.However, in the 1980s, a breakthrough in semen sexing technology was made by USDA researchers. The patents for this technology were subsequently licensed to a company named XY Inc. of Fort Collins, Colorado. Commercialization of sexed semen in the United States was initiated with a 2003 license granted to Sexing Technologies (STs) in Navasota, Texas. Sexed bovine semen is now commercially available.
Briefly, the method utilizes a flow-cytometer and associated nontoxic DNA dyes to sort sperm and simultaneously detect the 3-4% difference in DNA content between male and female sperm. The first step in this procedure is to dilute sperm to a very low concentration and stain them with a fluorescent dye. The sample is passed under pressure through the flow cytometer. As single sperm cells pass through an internal laser beam, the fluorescent dye is excited. Because of the larger X chromosome, female sperm emit less fluorescence than the Y chromosome bearing male spermatozoa. Detectors measure the amount of fluorescence and assign positive or negative charges to each droplet containing a single sperm. Charged deflector plates then split the single stream into three streams: positively charged particles containing one sex go one way and negatively charged droplets containing the other sex are deflected in the opposite direction, while uncharged droplets containing multiple sperm or unidentified sperm pass through the machine without being deflected. Based on multiple research trials, the procedure separates sperm of the two sexes with approximately 90% accuracy.
There are, however, limitations that have hampered widespread implementation of AI using sex- sorted semen. Reduced conception rate is an issue. Given the necessary manipulation, the sorting process negatively impacts sperm viability and longevity compared with normally cryopreserved semen (Weigel, 2004).
In addition, the procedure is extremely slow and relatively inefficient. To properly sort, sperm must be precisely oriented as they pass through the laser and fluorescence detectors in the flow cytometer. Due to the flat shape of bovine sperm heads, only about 30% are correctly oriented and half of these are female. This means only 15% of the sperm entering the procedure are recovered as a marketable product. Although 3000-5000 sperm of each sex can be sorted per second, it still takes approximately 1.3 hours of sorting to produce enough semen for a standard 20 million sperm/straw dosage. This resulted in commercialization of a product with about 10-fold fewer spermatozoa per insemination. Because of the low sperm numbers per dose and compromised sperm viability, sex-sorted semen is only recommended for use in well-managed, highly fertile virgin heifers. Data summarized to this time indicate that herds that typically achieve a 60-65% conception rate in virgin heifers with normal semen can expect a 45-55% conception rate with sexed semen. Sex-sorted semen is not currently recommended for use in lactating cows (because of inherently reduced conception rates, compared with heifers).
Cloning
Cloning is the creation of a new individual that is genetically identical to the source individual. Identical (monozygotic) twins come from an embryo that spontaneously splits, resulting in natural cloning. Cloning animals is a technique that may well assist animal industries in maintaining high quality livestock to supply future food and fiber needs. Identifying and reproducing superior livestock genetics ensures that herds are maintained at the highest quality possible. Animal cloning offers potential benefits to consumers, farmers, and endangered species. For example, it allows farmers and ranchers to accelerate the reproduction of their most productive and healthiest livestock. Cloning can be used to protect endangered species. For example, in China, panda cells are being kept on reserve should this species' numbers be threatened by extinction.
In 1997, cloning was revolutionized when Dr. Ian Wilmut and his colleagues at the Roslin Institute in Edinburgh, Scotland, successfully cloned a sheep named Dolly. Dolly was the first mammal cloned using nonembryonic cells. Wilmut and his colleagues transplanted a nucleus from a mammary gland fibroblast of a Finn Dorsett sheep into the enucleated egg of a Scottish blackface ewe. The nucleus-egg combination was stimulated with electricity to fuse the two and to stimulate cell division. The new cell divided and was placed in the uterus of a blackface ewe to develop. Dolly was subsequently born. Dolly was shown to be genetically identical to the Finn Dorsett mammary cells and not to the blackface ewe. This clearly demonstrated that she was a successful clone (it took 276 attempts before the experiment was successful). Dolly has since grown and reproduced several offspring of her own through normal sexual means. Therefore, Dolly is a viable, healthy clone.
Nuclear transfer in livestock using cells of the interstitial cell mass (ICM) of a donor blastocyst is relatively routine. Nuclear transfer is accomplished by the fusion of a donor cell to the unfertilized ova, which has been enucleated. Before the success of somatic cell nuclear transfer, it was possible to obtain cells from transgenic early bovine embryos as a source of nuclear material for donation to an enucleated oocyte.
The most common cloning method, known as "somatic cell nuclear transfer" or simply "nuclear transfer," requires two kinds of cells. One is a somatic cell, which is collected from the animal scheduled to be cloned (known as the "genetic donor"). A somatic cell is any cell other than a sperm cell or egg cell. The other kind of cell required for cloning is an egg cell, which is collected from a female of the same species. In the lab, the nucleus of the egg cell is removed. The nucleus from the donor somatic is then inserted into the enucleated egg. A charge is applied resulting in the fused egg.
The activated egg is then placed in a culture medium. Over the course of several days, a blastocyst forms. The blastocyst is subsequently implanted in the uterus of a recipient female (sometimes referred to as "surrogate mother") where it continues to develop. After a full-term pregnancy, the recipient gives birth to an animal that is essentially the identical twin of the genetic donor. Since Dolly, several university laboratories and companies have used various modifications of the nuclear transfer technique to produce cloned mammals, including cows, pigs, monkeys, and mice. This technique is outlined in Figure 19.26.
Transgenic animals
Farmers began manipulating genome of animals soon after domestication of livestock began. Certainly most
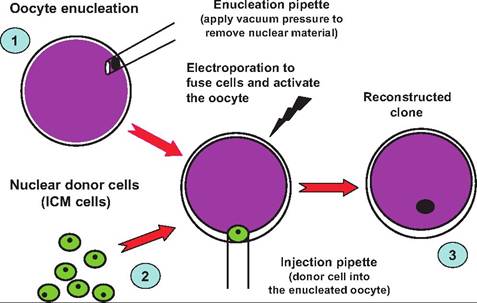
Fig. 19.26. Overview of micromanipulation and cloning. Step 1 indicates the removal or enucleation of the existing nucleus of a fertilized oocyte. This is accomplished by micromanipulating the egg under differential contrast or phase contrast microscope. The egg is held with slight vacuum with a microscopic blunted "holding" pipette (not shown) while a fine dissection pipette is used to aspirate the nucleus of the egg. Step 2 indicates the collection of the nucleus of a donor cell, in this case cells from the interstitial cell mass (ICM) from the embryo of an animal scheduled for cloning. In step 3, the donor cell nucleus is injected, and a slight electrical charge is used to induce fusion of the donor nucleus and egg. The newly generated egg is allowed to develop, then frozen or implanted in a surrogate dam.
of this genetic gain arose from the selection of individuals considered superior to be retained as breeding stock. Production of transgenic animals is in many ways an extension of this effort. This became possible as a consequence of the ability of scientists to develop recombinant DNA and thereby isolate individual genes for the transfer of copies into the genome of other animals. A "transgenic" animal is one that integrates recombinant DNA into its own genetic material.
The primary method for the production of transgenic animals is through the introduction of foreign genes by microinjection of DNA into one pronucleus of fertilized ova. This procedure was described in 1980 for mice and has been used mostly for the production of transgenic animals. Fertilized ova can be collected prior to the time of cell division after superovulation or following IVF of oocytes collected from slaughterhouse ovaries. The apparent potential viability of in vivo collected zygotes is higher than for the superovulated zygotes, but the number of potential cells available for gene insertion makes the slaughterhouse-provided oocytes more practical for transgenic work in cattle. Review summaries indicate that for DNA microinjected into over 11,500 IVF slaughterhouse-derived ova, only 9% developed to the morula/blastocyst stage. Of 478 embryos transferred to recipients, 90 (19%) produced calves. Ten percent of the calves were transgenic and a heifer from the study was induced to lactate and shown to secrete human Oc-Iactalbumin in her milk. The rate of transgenesis, based on microinjected zygotes, was.08% (Ayares, 2000). This technique is illustrated in Figure 19.27.
Indeed, a variety of proteins used in human medicine (human growth hormone, insulin, erythropoietin, etc.) are recombinant proteins derived from the transfer of genes coding for the production of these proteins
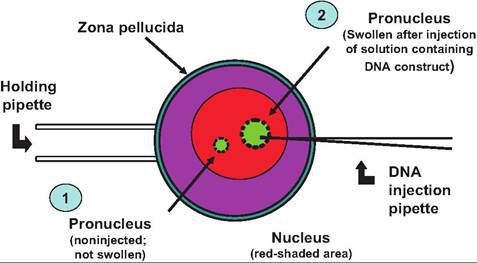
Fig. 19.27. Microinjection and transgenic animals. Step 1 indicates the normal appearance of the pronucleus of a fertilized egg. For insertion of the gene constructs, the egg is positioned with a holding pipette and the injection pipette containing a solution with multiple copies of the DNA construct (step 2) is used to inject a small volume (~1 pl) into the pronucleus. In response, the organelle swells.
into bacterial cells. Bacterial cells containing these choice proteins are subsequently grown in large incubators and the proteins harvested and purified. The recombinant bovine somatotropin used to enhance lactation performance in dairy cows, the rennet used in cheese making, and much of the insulin used to treat human diabetics is derived in a similar process.
In recent years, scientists have utilized these technologies to link genes of interest with gene promoters that would drive the production of specific proteins in selected tissues and especially the mammary gland. It has been possible to utilize promoter regions of specific milk protein genes (caseins, Oc-Iactalbumin, β-lactoglobulin) to direct secretion of a variety of proteins into milk. These transgenic animals are then bred, and the lactating animals are used as living bioreactors, for example, the milk used to purify and isolate the desired protein products (Table 19.3).
The production of pharmaceutical proteins (assuming investment and support continues) likely holds a more immediate application than transgenic tools to improve livestock performance. Safe and effective proteins produced from transgenic animals can be produced with current technologies and products could be available in unlimited quantities and at reasonable costs to consumers. Many of these products may be the result of using the mammary glands of sheep and goats and possibly cows and pigs as bioreactors for the production of human recombinant proteins.
Despite the potential benefits, there are limitations in technology because of availability of appropriate genes for DNA insertion and the inefficiencies of production of transgenic livestock. More genes are likely to become available as more basic genome projects identify candidates. But low efficiencies for transgene integration and low embryonic survival rates because of embryo manipulation are large impediments. Integration efficiencies are less than 30% in laboratory mice, and lower in livestock species. While the low integration and reproductive efficiencies are acceptable for laboratory species, these costs become less acceptable for use in livestock species.
Choice of gene construct is probably the most important decision for attempting to design livestock to have a significant impact on the agricultural and consuming community. The construct usually contains a regulatory element (promoter, enhancer) from one gene ligated to a structural gene sequence. This combination allows for theoretical direction of the expression of the structural gene to specific areas of production, at specific times, and allows for regulation of the amount of product produced.
Transgenic technology for the improvement of animal production or performance has been touted for nearly two decades since the first publication of the "super" mouse with an added growth hormone (GH) construct. The prospect for improvement of dairy, beef, swine, and sheep production and health through transgenesis is still rudimentary. Part of the delay in real progress with transgenic animal production has been the fact that DNA injected into the egg (with current techniques for livestock species) is incorporated at random sites in the genome. This can produce negative effects. For example, addition of a GH gene in swine lead to overproduction of GH and insulin-like growth factor I (IGF-I) and subsequent physiological conditions similar to excessive GH production in acromegaly. Additionally, the high cost of animals and their care, the lack of inbred lines, long generation intervals, and production of small numbers (1 or 2) of offspring per female in sheep and cattle have lowered the chances for meaningful progress in livestock species with this approach. However, as indicated previously, utilization of the mammary gland as a bioreactor offers interesting possibilities for large scale production of recombinant proteins that cannot be done with bacterial cell cultures.
Table 19.3. Selected listing of milk protein gene promoters and expression of recombinant proteins in milk.
| Promoter and Expressed Protein | Animal | Concentration per Milliliter |
| Bovine Oi-Iactalbumin | Rat | 2.4mg |
| Bovine Oi-Iactalbumin | Mouse | 1.5 mg |
| Human Oi-Iactalbumin | Cow | 2.4mg |
| Bovine αs1-casein hEPO1 | Mouse | 0.2 mg |
| ItIGF-I2 | Rabbit | 1mg |
| IiLactoferrin | Cow | Not reported |
| Goat β-casein | Mouse | 22 mg |
| IiLAtPA3 | Goat | |
| hAnti-thrombin III | Goat | |
| Ovine β-lactoglobulin ItAAT4 | Mouse | 5 μg to 21 mg |
| ItAAT | Sheep | 5μg to 35 mg |
| IiFactor IX | Sheep | 25 ng |
| Iilnterferon γ | Mouse | 2θ ng |
| ItFibrinogen | Sheep | 5 mg |
| Mouse whey acidic protein ItGH5 | bgcolor=white>Mouse 3.5 mg | |
| Protein C | Mouse | 3μg |
| Protein C | Pig | 1mg |
| ItFactor Vlll | Pig | 2.7mg |
1hEPO-recombinant human erythropoeitin.
2hlGF-l-recombinant human insulin-like growth factor-l.
3hLAtPA-recombinant human long-acting tissue plasminogen activator.
4hAAT-recombinant human α-1-antitrypsin.
5hGH-recombinant human growth hormone.
Adapted from Gwazdauskas (2002).
Chapter summary
In animal agriculture, reproductive success is required at some point along the production chain. Population biologists describe two general reproductive patterns: r-selection and K-selection. The r-selection depends on production of many offspring but minimal or no parental care once the animals are hatched or born. Examples include amphibians or in aquaculture among agricultural industries. The K-selection is more typical, where parental investment is substantial because the number of offspring is low, with 1-2 offspring or small litters or clutches. Expenditures can be dramatic in terms of time (behavioral, training, protection) and direct energy expenditures (lactation and nursing, gathering/hunting). Animal husbandry and domestication of animals evolved to improve the odds of reproductive success as well as improve efficiency of growth and development of offspring.
Female reproductive system
The structures of the female reproductive tract include the ovaries, oviducts, uterus, cervix, vagina, and external genitalia. In most farm animals the reproductive tract is positioned below the rectum. For cows and mares, this allows the producer to manually evaluate the reproductive tract by manipulation per rectum. This allows determining functional status of the ovary, diagnosis of pregnancy, or manipulation of the tract for Al.
The histology of the reproductive tract reflects function of the various segments. Similar to the GI tract, the outside is covered by serosa of simple squamous epithelial cells, followed by the muscularis with layers of longitudinal and circular smooth muscle, the submucosa with associated blood and lymph vessels and nerves, and finally the mucosa. The types of epithelial cells lining the mucosal surface vary by location along the tract. For example, the vaginal epithelium is stratified squamous but the relative thickness of this layer varies during the estrus cycle (thicker during the follicular phase and thinner during the luteal phase). The uterus is lined by simple cuboidal epithelial cells but there are also many uterine glands that extend into the underlying submucosa. Also, compared with other regions, the muscularis is markedly more extensive. Like the vaginal lining, the activity of the uterine glands and the surface cells varies during the estrus cycle and particularly during pregnancy. The epithelial cells of the oviduct are highly specialized columnar epithelial cells with cilia. Secretions and movement of the cilia aid the movement of spermatozoa such that fertilization of the egg transiting from the ovary usually occurs in the ampulla region of the oviduct. When conditions are suitable, the fertilized ovum passes into the uterus where implantation occurs. The ovaries, analogous to the testes in the male, are the primary reproductive organs since they produce the female gametes (singular = ovum; plural = ova).
The ovaries are paired structures located at the ends of the uterine horns. In postpubertal animals the ovary undergoes dramatic cyclic development. Ovulation is the rupture of the mature follicle and the release of the ovum. It is picked up by the infundibular (funnel-like) end of the oviduct. The remaining ruptured follicle(s) are transformed into corpus Iuteum (singular) or corpora Iutea (plural) that produce large quantities of progesterone. If fertilization does not occur, the corpus Iuteum regresses and a new crop of follicles mature.
Newly recruited follicles produce estrogen and selected follicle(s) proceed to undergo ovulation. The ovary is oval to round in shape with distinct regional differences and blister-like structures—the follicles— near the outer surface. Instead, beneath the outer protective tunica albuginea, within the ovarian cortex,
there are populations of oocytes, which are recruited to develop into the mature follicles. The ovarian cortex also houses the corpus Iuteum as well as older degenerated corpora Iutea called corpora albicantia. The very center of the ovary is called the ovarian medulla and it contains the blood, nerves, and lymphatic vessels that supply the ovary.
Grossly, the uterus consists of a body, a cervix (or neck), and two uterine horns. However, there are variations in the shape of the uterine body and arrangement of the horns. Three distinct types of uteri are common: simplex, bipartite, and bicornuate. Primates have a simplex uterus. There is a large uterine body but essentially no uterine horns. The bicornuate uterus is characterized by the presence of two very distinct uterine horns and a relatively small heart-shaped uterine body. Examples include the sow, rats, and mice. In the bipartite, the uterus appears nearly divided into two parts nearly to the base with distinct uterine horns (mare, cow, and ewe).
The internal lining (endometrium) of the uterus varies markedly during the estrous or menstrual cycles and during pregnancy. The tissue is highly glandular with an epithelial surface of simple columnar epithelium in the mare but stratified columnar epithelial cells in ruminants. Simple branched tubular glands provide secretions—called uterine milk—that are especially important during estrus and pregnancy. In many animals these uterine glands are scattered throughout the endometrium. But in ruminants, the internal uterine surface is punctuated by caruncles that are not glandular. These mushroom-like caruncles provide sites for attachment of fetal membranes in these animals. The smooth muscle of the uterus (the muscularis) is frequently called the myometrium.
Estrous cycles and ovulation
The period when the female is receptive to sexual activity is called estrus or, more commonly, heat. An estrous cycle is simply the time from the beginning of one estrus period to another. Estrus is a noun. (A cow displays estrus or heat). The average length of an estrous cycle in the Holstein dairy cow is 21 days. Animals that exhibit only one estrous cycle per year are monoestrous. The estrous period lasts for several days. Examples are dogs, wolves, foxes, and bears. In contrast, polyestrous species exhibit multiple cycles each year. During periods of inactivity the animals are as described anestrous. These periods may be seasonally induced or related to physiological state, for example, early postpartum anestrous in suckling beef cows.
With the dairy cow as a model, tonic secretion of FSH and LH cause a cohort of primary follicles to enlarge and become small antral (fluid filled) follicles. These follicles provide those that are eligible for activation during sequential estrous cycles. The estrous cycle is divided into two phases, named after the dominant structures that are present on the ovary. The follicular phase is relatively short, -20% of the estrous period, and the preovulatory follicles that produce estradiol are in control. The follicular phase encompasses the time from the regression of the corpora Iutea to the time of ovulation. Several waves of follicular development occur during the estrous cycle in cattle. Beginning after ovulation, groups or clusters of small or medium antral follicles become especially sensitive to gonadotropins and are recruited. Among these follicles, several are selected and begin to mature. However, typically only one of the selected follicles will win the maturity race to become the dominant follicle. Other follicles in the class undergo atresia (regression). Even the dominant follicle is destined to undergo atresia if it is so unlucky to have been recruited in the first or second wave of follicular development during the estrous cycle.
The LH surge and ovulation in most farm animals (cow, sow, ewe, and mare) take place regularly, independent of copulation. These animals are spontaneous ovulators. In other animals, such as rabbits, mink, camels, llamas, and alpacas, ovulation requires copulation (induced ovulators). After ovulation, granulosa cells lining the now-empty follicular cavity divide and undergo luteinization in response to high levels of LH. This results in the appearance of the corpus luteum. The CL is a powerful endocrine tissue that produces large amounts of progesterone, which prepares the uterus to receive the ovulated ovum or ova. If fertilization and implantation of the ova is successful, this produces maternal recognition of pregnancy and the usual regression of the CL does not take place. If pregnancy is not established, the CL regresses and the cycle repeats.
In females, the neurons of the hypothalamus that secrete GnRH must acquire the ability to secrete enough GnRH in response to feedback from ovarian estrogen to stimulate ovulation. This depends on interactions between hypothalamic hormones, anterior pituitary hormones, and especially the impact of gonadotropin-releasing hormone or GnRH on secretion of FSH and LH.
Secretion of GnRH is controlled by a tonically acting center located in the area of the dorsomedial nucleus of the hypothalamus. Before puberty, secretion of GnRH occurs at a relatively slow frequency and the amplitude of the each of the secretory events is also rather low. As ovarian development progresses, the secretion of estradiol from waves of growing follicles also increases. Over time, as puberty approaches, the estradiol (along with effects of environment, nutrients, and social interactions) has an escalating impact to increase the frequency and amplitude of bursts of GnRH secretion from the tonic control center. This produces more FSH and LH and, in a positive cascade, more follicular activity.
However, ovulation requires not just low-level secretion of LH but rather a marked surge in the concentration. This is called the preovulatory LH surge. This depends on the activation of a second population of hypothalamic nuclei called the surge center. These nuclei are located more anteriorly in the preoptic and anterior hypothalamic nuclei. The prepubertal female is characterized by having insufficient ovarian-derived estradiol to stimulate the surge center and reduced sensitivity to estradiol. As she matures, her hypothalamus becomes progressively more sensitive to estradiol, so that cycles of ovulation then begin to occur.
Fertilization and pregnancy
After a successful ovulation and insemination, spermatozoa ascend through the cervix, uterine body, and the oviduct. But for fertilization to succeed, the spermatozoa must undergo a process called capaci- tation. These are biochemical changes induced by secretions of the female reproductive tract. Fertilization typically occurs when the oocyte and spermatozoa meet in the ampulla region of the oviduct.
The embryo develops into a blastocyst, hatches from the surrounding zona pellucida, develops a functional trophoblast, and secretes signals that allow maintenance of the corpus luteum. After fusion of the male and female pronuclei, the fertilized egg is called a zygote. By contrast, a fetus—a potential offspring still within the uterus—can generally be recognized as a member of the species. Soon after syngamy the zygote initiates a series of mitotic divisions. Each of the cells at this time is called a blastomere. Subsequent divisions produce 4, 8, and 16 identical daughter cells. At these early stages the blastomeres are totipotent. In other words, each of the individual cells is capable of giving rise to a fully formed offspring. For example, identical twins can be produced in experimental situations by separating individual blastomeres, placing then into surrogate zona pellucida, and allowing development in the uterus of a recipient female. Once beyond the 16-cell stage it becomes impossible to accurately count the growing ball of cells, so the structure is called a morula. The blastocyst develops and is characterized by the presence of inner cell mass (ICM), a fluid-filled cavity called a blastocoele, and a layer of cells around the periphery called the trophoblast. The ICM develops into the embryo while the trophoblast gives rise to the fetal component of the placenta.
Implantation is the attachment of the free-floating blastocyst to the uterine epithelium and growth or penetration of the epithelium by embryonic tissues. Just how dramatic and extensive this interaction is varies between species. Placentation is development of the extraembyronic membranes or the placenta. The placenta allows for exchanges between the maternal and fetal circulations so that nutrients can be supplied and wastes removed. The chorion is the outer membrane and therefore is in contact with the maternal uterine wall. The next layer toward the fetus is the allantois. The amnion is the closest membrane to the fetus. It also forms a fluid-filled cavity in direct contact with the fetus. When parturition occurs, the allantoic sac is expelled, followed by the amniotic fluid.
The placentation of most farm animals is classified as epitheliochorial. This means that the chorion of the fetus is in direct contact with the uterine surface. This is the least intimate arrangement of the types of placentas. In hemochorial placentation, fetal vessels and chorion are invaginated so that there is direct contact between a single layer of chorion epithelial cells and maternal blood. In hemoendothelial placentation, maternal blood can directly bathe the outer surface of chorionic capillaries. The number of layers separating fetal and maternal circulations is important in part because this markedly affects transfer of immunoglobulins in utero. Since most farm animals have epitheliochorial connection, calves, for example, are born without protective antibodies from the dam.
Placentation patterns also vary at a gross anatomy level based on the location or distribution of sites for exchange between the placental and the uterus. In the horse and pig, extensions from the chorion (chorionic villi) project into crypts scattered over the entire surface of the endometrium. This is called a diffuse type of placenta. Ruminants have scattered attachments, which depend on a cotyledonary type of connection. Exchange takes place at distinct structures called placentomes. The placentomes are formed by the combined invagination of specific regions of the chorionic membrane, the cotyledons, into the button-like projections from the surface of the endometrium, the caruncles. Caruncles project from the surface of the uterus. Zonary placentas
(dogs and cats) are characterized by a major region of exchange that forms in an area near the center of the developing conceptus essentially in a band.
Parturition occurs in three phases: (1) initiation of uterine contractions, (2) expulsion of the fetus, and (3) expulsion of the fetal membranes. During gestation high progesterone concentrations maintain a "quiet" uterus; as birth approaches, Iuteolysis of the dominant CL and/or decreasing placental production of progesterone couples with increasing estrogen concentrations to prepare for parturition. Increasing estrogen stimulates expression of oxytocin receptors in the myometrium. Activation of the fetal pituitary-adrenal axis is essential for the initiation ofparturition. The fetus becomes "stressed" and secretes ACTH and then glucocorticoids. High concentrations act to remove the progesterone block by stimulation of placenta enzymes that increase the conversion of progesterone to estradiol. Fetal corticoids promote synthesis of PGF2oc, which also acts to remove the progesterone block.
Neural signals from the cervix induce oxytocin and release more contractions, more pressure, and more oxytocin in a positive cascade. As the fetus enters the cervical canal, the first stage of parturition is complete. With the fetus positioned, the feet and head increase pressure on the fetal membranes, which rupture, releasing amniotic fluid. As the fetus passes the birth canal it becomes hypoxic. This promotes fetal movement and more uterine contraction.
Male reproductive system
The male reproductive tract consists of the testis, epididymis, spermatic cord, accessory sex glands, and the penis. The testes produce spermatozoa and testosterone. The epididymis provides the environment for spermatozoa to mature and be stored. The accessory sex glands produce seminal plasma and fluids and the penis is the structure needed for copulation. Male birds have paired abdominal testes located anterior to the cranial lobe of each kidney. The vas deferens emerges medially and passes cau- dally alongside the ureters to the cloaca, where it has a common opening with the ureter in the urodeum. Few male birds have a penis-like structure. Fertilization is achieved by pressing the cloaca of the male and female together. This involves positioning the male on the back of the female with a twisting of his tail under hers and the touching of the cloaca.
Testicular parenchyma includes the seminiferous tubules and the interstitial cells of Leydig. The seminiferous tubules are the key to spermatogenesis. The end of each seminiferous tubule joins with the rete tubules that are continuous with the epididymis. Sertoli cells are the only somatic cells that are part of the internal structure (inside the basement membrane) of the seminiferous tubules. Each of the Sertoli cells tends or anchors a maximum number of germ cells. Sertoli cells are analogous to the granulosa cells of the ovarian follicles. The basal region houses the spermatogonia and the early spermatocytes. The adluminal region has all the other germ cells, that is, primary and secondary spermatocytes, spermatids, elongated spermatids, and mature spermatozoa.
Reproductive technologies
A variety of techniques, often called ART, for example, IVF, superovulation, ET, and freezing, are popularized because they are used to assist couples incapable of conceiving naturally. However, these tools have their origins in reproductive physiology of laboratory and farm animals. For example, scientists worked for many years to perfect IVF before it was recognized that sperm cells have to undergo Capacitation in the female reproductive tract before fertilization. This ultimately led to methods to induce Capacitation, and, therefore, routinely successful IVE Advances in one aspect of reproduction thus allow progress in other areas. For example, ET would not be practical without endocrine tools to induce superovulation and methods to routinely collect, fertilize, and freeze eggs. Cloning and development of transgenic animals depended on development of techniques to visualize, micromanipulate, and culture eggs and embryos.
Review questions and answers are available online.
References
Ayares, D.L. 2000. Transgenic protein production: achievements using microinjection technology and the promise of nuclear transfer. J. Anim. Sci. 78(Suppl. 3): 8-18.
Bearden, HJ., J.W. Fuquay, and S.T. Willard. 2004. Applied Animal Reproduction. Pearson Prentice Hall, Upper Saddle River, New Jersey.
Frandson, R.D., W.L. Wilke, and A.D. Fails. 2003. Anatomy and Physiology of Farm Animals, 6th Edition. Lippincott Williams and Wilkins, Ambler, Pennsylvania.
Gwazdauskas, EC. 2002. Gamete and embryo technology- transgenic animals. In Encyclopedia of Dairy Science. Edited by H. Roginski, JW. Fuquay, and P.F. Fox. Academic Press, London.
Krisherz R.L.z J.R. Gibbons, and F.C. Gwazdauskas. 1995. Nuclear transfer in the bovine using microinjected donor embryos: assessment of development and deoxyribonucleic acid detection frequency. J. Dairy Sci. 78: 1282-1288.
Peeler, I.D.z R.L. Nebelz R.E. Pearson, W.S. Sweckerz and A. Garcia. 2004. Pregnancy rates after timed AI of heifers following removal of intravaginal progesterone inserts. J. Dairy Sei. 87: 2868-2873.
Perry, G.A. 2012. Harnessingbasic knowledge of factors controlling puberty to improve synchronization of estrus and fertility in heifers. J. Anim. Sei. 90: 1172-1182.
Randall, D., W. Burggen, and K. French. 2002. EckertAnimal Physiology. W. H. Freeman and Company, New York.
Sakaguchi, M. 2011. Practical aspects of the fertility of dairy cattle. J. Reprod. Dev. 57: 17-33.
Senger, P.L. 2003. Pathways to Pregnancy and Parturition. Current Concepts, Inc., Pullman, Washington.
Weigel, K. A. 2004. Exploring the role of sexed semen in dairy production systems. J. Dairy Sei. 87: E120-E130.
Whittow, G.C., ed. 2000. Sturkie's Avian Physiology, 5th edition. Academic Press, San Diego, California.
Wiltbank, M.C., R. Sartori, M.M. Herlihy, J.L.M. Vasconcelos, A.B. Nascimento, A.H. Souza, A.P. Cunha, A. Keskin, J.N. Guenther, and A. Gumen. 2011. Managing the dominant follicle in lactating dairy cows. Theriogenology 76: 1568-1582.