ROE DEER AND OTHER UNGULATE PAPILLOMAVIRUSES
KAROLY ERD ELYI
Central Agriculture Office, Veterinary Diagnostic Directorate, Budapest, Hungary
Infectious Diseases of Wild Mammals and Birds in Europe, First Edition. Edited by Dolores Gavier-Widen, J.
Paul Duff, and Anna Meredith. © 2012 Blackwell Publishing Ltd. Published 2012 by Blackwell Publishing Ltd.Deer fibropapillomatosis or deer fibromatosis is a cutaneous benign neoplastic disease occurring in a wide range of cervid species, caused by deltapapillomaviruses of cervids.
Ruminants, including many deer species, are infected with the second most numerous and diverse range of papillomaviruses after humans. Of the three artiodactyl-specific papillomavirus genera, deltapapillomaviruses infect the widest host range, encompassing ovine, bovine and several cervid species.
AETIOLOGY
Papillomaviruses have been identified in skin tumours of European moose (Alces alces), reindeer (Rangifer tarandus), roe deer (Capreolus capreolus), red deer (Cervus elaphus) and North American deer species (e.g. white-tailed deer). A common feature of these viruses is that they induce the development of skin tumours with a marked connective tissue component. Cervid papillomaviruses causing cutaneous fibromatosis or fibropapillomatosis share this genus with Bovine papillomavirus 1 and 2 (BPV1 and BPV2) and Ovine papillomavirus 1 and 2 (OaPV1 and OaPV2).
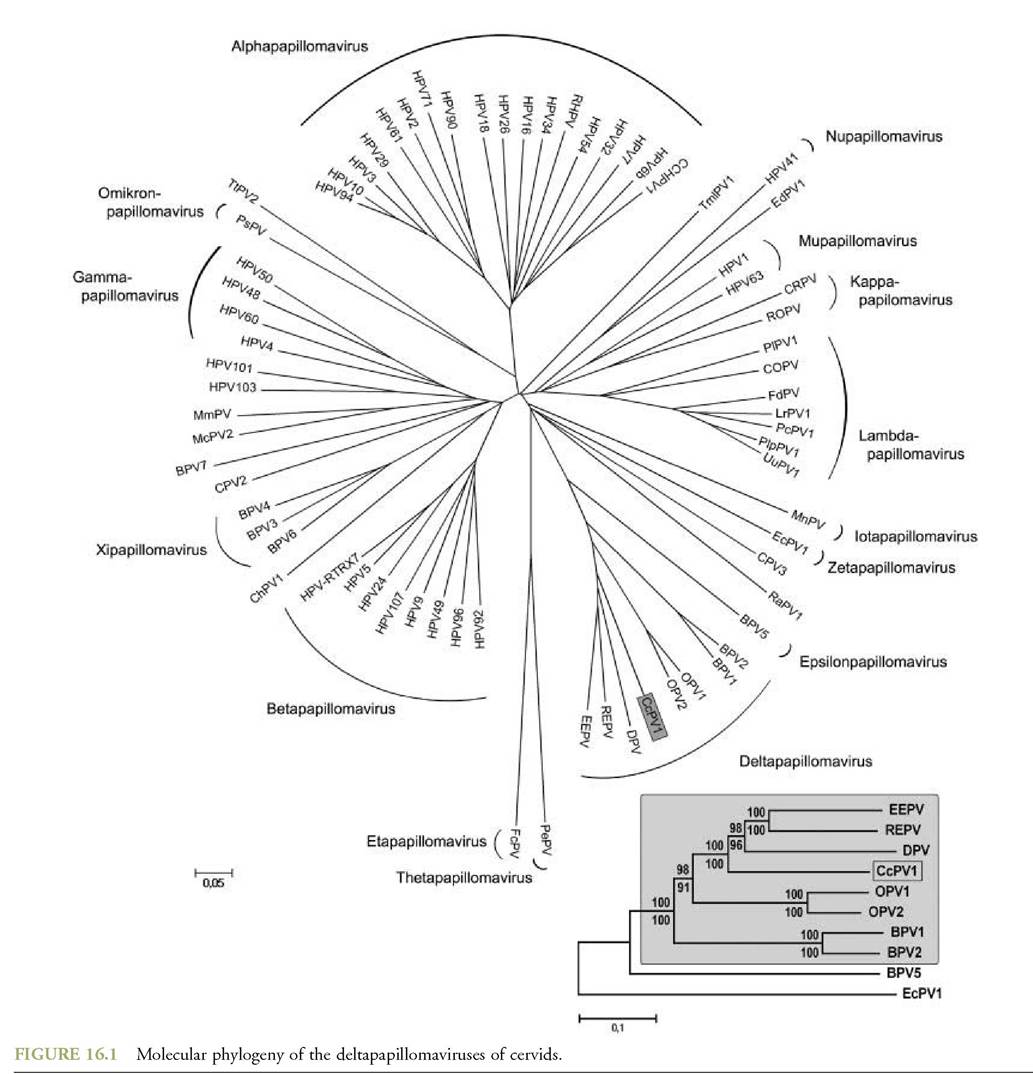
The genomes of the following deltapapillomaviruses of cervids have been sequenced, characterized and deposited in GenBank: Deer (White-tailed deer — Odocoileus virgin- ianus) papillomavirus (OvPVl), Reindeer papillomavirus (RtPV1), European moose papillomavirus (AaPV1) and Western roe deer papillomavirus (CcaPVl). In addition to the above, characterization of a sequence belonging to the Red deer papillomavirus shows that it is also a deltapapillomavirus, very closely related to the Roe deer virus^x∖ Earlier Southern blot analyses(2) have already indicated this relationship by establishing that Red deer papillomavirus genomic DNA cross-hybridizes with BPV1/BPV2 DNA (Figure 16.1).
The size of the non-enveloped, icosahedral deltapapillomavirus capsid is approximately 55 nm in diameter, it is constituted from 72 capsomeres and contains the doublestranded, circular DNA genome of 8—8.4kbp. All cervid papillomaviruses exhibit homologous genetic structures containing a total of nine open reading frames (ORF) coding early (E) regulatory proteins responsible for replication and transforming properties, the late (L) structural capsid proteins and the ubiquitous, non-coding long regulatory region (LRR). The major capsid protein is the main immunogenic epitope of papillomaviruses. The main oncogene of deltapapillomaviruses is the E5 ORF, but the additional fibroblastic properties typically expressed in deer fibromas may actually be the effect of the E9 ORF identified in all cervid deltapapillomaviruses
The current classification of papillomaviruses is based on the comparison of complete L1 ORF nucleotide sequences. Distinct papillomavirus genera share less than 60% L1 ORF nucleotide sequence identity, whereas papillomavirus ‘species’ within a genus share approximately 60—70% nucleotide identity.
EPIDEMIOLOGY
Skin lesions caused by papillomaviruses have been described in cervid species from both Europe and North America. In Europe, endemic infections typically occur in roe deer populations of Central-Eastern Europe, i.e. Hungary and neighbouring areas(3) and in European moose throughout Sweden, with high frequency in the middle of the country. Papillomavirus-induced fibropapillomas also occur sporadically in red deer throughout Europe and in reindeer in Sweden (Figure 16.2).
Cutaneous fibromas and fibropapillomas of roe deer were observed in parts of Hungary from the early 1960s, and they received more attention 30 years later because of the allegedly growing incidence of this endemic disease. The prevalence of the infection in endemic areas was found to be between 0.2 and 1.1%, which is similar to the prevalence of OvPV1-induced fibropapillomatosis in North American cervids.
A similar 1.1% general prevalence of fibromatosis has been observed in moose in Sweden, but prevalences as high as 50% have been reported from certain areas.Fibropapillomas are most commonly seen in mature roe deer and European moose — unlike red deer fibropapillo- mas, which are mostly found in young animals, similar to OvPV1 infection of white tailed deer. Males and females of both roe deer and European moose are equally affected by fibromatosis, but the prevalence is much higher in white-tailed deer bucks in North America.
Papillomaviruses are highly host-specific. The endemic nature of both roe deer and European moose (also named European elk) fibropapillomatosis suggests the possibility that additional, genetically determined susceptibility factors of the hosts may have contributed to the evolution of these diseases within certain subpopulations of each species. In the case of CcaPV1, this possibility is supported
by the genetic differentiation pattern of European roe deer populations, as the boundary between the two major genetic clusters of European roe deer (southern and eastern European versus western, central and northern European) corresponds to the westernmost distribution limits of roe deer fibropapillomatosis(4).
The exact method of cervid papillomavirus transmission in nature are not known. The primary transmission route of cutaneous papillomaviruses in general is through direct contact with a contaminated environment or infected animals, which shed the virus from the epithelial surface of tumours. As viral particles have to penetrate the
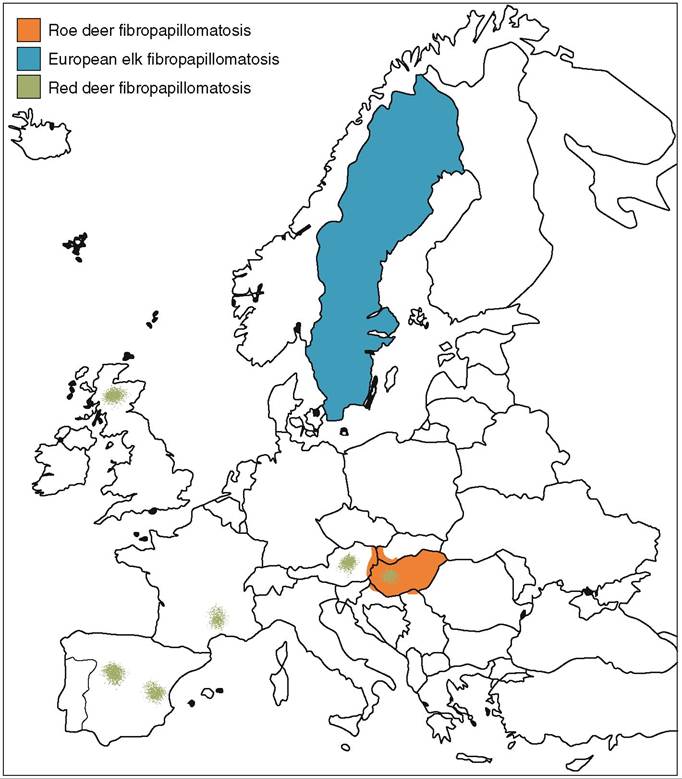
FIGURE 16.2 Distribution map of deltapapillomaviruses of cervids in Europe.
superficial epidermis to cause infection, it is assumed that infection is acquired predominantly through superficial skin lesions.
Several aspects of deer behaviour, such as group- formation, scent marking, rubbing of antlers on trees during velvet shedding and fighting, may contribute to transmission. The common localization of lesions on the head, neck, extremities and belly supports these possibilities.It has been suggested that arthropod vectors may also play a part in the epidemiology of deer fibromatosis. Circumstantial evidence from an epidemiological study of roe deer fibropapillomatosis does not rule out the potential role of arthropods in papillomavirus transmission1-3). The spatial distribution of the disease was correlated with the presence of waterways, habitats especially suitable for arthropod vectors, independently of host density. Alternatively, this correlation could also be the potential effect of seasonal roe deer aggregation in these habitats. The arthropod transmission hypothesis still requires confirmation.
Pathogenesis, pathology and immunity
All deltapapillomaviruses induce the development of cutaneous fibropapillomas and fibromas(5), and OvPV1 and AaPV1 infections are also associated with sporadic cases of nodular pulmonary fibromatosis(6). An active papillomavirus infection is established when viral particles enter basal epithelial cells. Viral genomes are present in low numbers within the nuclei of basal epithelial cells (keratinocytes) in episomal form and they maintain low- level replication, using the normal cell cycle. After the transcription of early regulatory genes, substantial papillomavirus genome replication and transcription only occurs in an advanced state of epithelial cell differentiation. This process culminates in capsid formation and virus assembly taking part in the stratum granulosum. Simultaneously with the epithelial changes, the proliferation of fibroblasts is induced in the underlying dermis, resulting in the development of the main tumour mass and the clinical manifestation of the disease.
Lesions induced by deltapapillomaviruses are limited almost exclusively to the epidermal squamous epithelium and the connective tissue of the dermis. Apart from the infrequent occurrence of lung fibromatosis, these viruses do not cause lesions in other organs. Deer fibromas and fibropapillomas do not undergo malignant transformation and the nodular lung lesions induced by AaPV1 and OvPV1 are not considered true tumour metastases, as they are probably the consequence of fibroblastic transformation induced by disseminated viral DNA.
Both the progression and regression phase of cutaneous fibropapillomatosis are protracted processes. Experimental infections with OvPV1 in white-tailed deer resulted in the development of fibropapillomas at the inoculation site within 2—3 weeks, and their regression within 7—10 weeks, post-infection. Natural infections with AaPV1 and CcaPVI are believed to have a course of several months or even years. Although the majority of papillomavirus infections induce certain levels of humoral immune response, cell-mediated immunity plays a major role in clearing the infection. Tumours get progressively infiltrated by lymphocytes and macrophages, and marked hyalinization is also observed during the regression process.
Fibropapillomas of roe deer are conspicuous skin tumours. They are firm, round, raised or pedunculous, most often multiple skin tumours of varying size, typically localized on the head, neck, belly and legs of infected animals (Figure 16.3). In European moose (elk), fibromas are found mainly on the head, neck, shoulders and sides and on the extremities.
Typical fibropapillomas of roe deer are firm, round and hairless, and covered with smooth or verrucous epidermis, but some tumour surfaces may exhibit rough, papillary structure. Tumours are often pigmented and the surface of
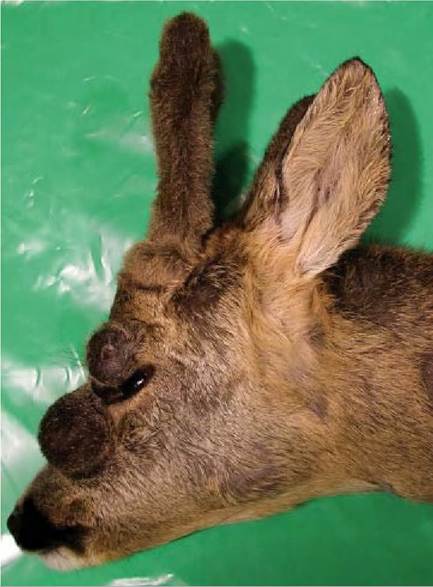
FIGURE 16.3 Macroscopic lesions of roe deer papillomatosis on the face.
larger lesions is frequently eroded and ulcerated. The cut surface of the tumour mass exhibits the compact, shiny, white appearance of firm, compact connective tissue. The lesions are on average 3—5 cm in diameter, but they can sometimes reach 11—17 cm. They most commonly appear on the head, neck, belly and legs of the infected animal. Although the number of fibropapillomas on one animal is usually less than 10, individuals carrying more than 150 tumours can often be found.
Microscopically the main tumour mass consists of groups of proliferating fibroblasts embedded in a mesh of collagen fibres, located in the stratum reticulare of the dermis. Tumours are covered with the looser connective tissue of the upper dermis and the epidermis, showing signs of acanthosis and hyperkeratosis. The cytoplasm of enlarged keratinocytes in the stratum spinosum and stratum granulosum contain aggregations of keratohyaline granules, and their nuclei are vacuolated.
Both papillomavirus L1 antigen and DNA can be visualized in the epidermis covering the tumour stroma. Papillomavirus DNA homologous to the L1 ORF sequence of CcaPV1 was found to be present in the nuclei of some stratum basale keratinocytes and in large numbers of stratum spinosum and stratum granulosum keratinocytes. Papillomavirus major capsid antigen can be found in both the cytoplasm and nuclei of keratinocytes of the stratum granulosum, but antigen aggregations representing complete papllomavirus particles are often observed between the keratin layers of the stratum corneum.
Nodular lung fibrosis occasionally associated with OvPV1 infection in European moose are multiple, firm, white nodules, 1—10 mm in diameter, formed by bundles of connective tissue located in the alveoli. The presence of AaPV1 DNA was demonstrated in these lesions by in situ hybridization, but virus particles were not detected.
CLINICAL SIGNS AND TREATMENT
In the majority of cases the presence of cutaneous fibromas and fibropapillomas does not have additional clinical effects and does not affect the general body condition, particularly in animals with few tumours. However, tumours on the head may obstruct vision or affect the ability of the animal to feed. This may eventually result in the decline of body condition and death of the infected animal due to emaciation. Tumours of extreme size may also hinder the normal movement of animals. It is not uncommon to find an extreme number of tumours on certain animals, coinciding with poor general condition. There is a possibility that these cases occur in immunosuppressed hosts or in animals simultaneously affected by other underlying illnesses.
Similar to the approach used in BPV infections of cattle, the treatment of infected captive deer might be attempted by auto-vaccines prepared from the lesions of the infected animal.
DIAGNOSIS
Papillomaviruses cannot be routinely propagated in vitro on tissue cultures. Diagnostic methods are therefore directed towards the demonstration of viral DNA or antigen in clinical or pathological samples. PCR can be used successfully for the demonstration of cervid papillo- maviruses(3’4). The method of rolling circle amplification (RCA) suitable for the detection of circular DNA is widely used for the detection of novel papillomaviruses and the amplification of complete genome sequences. Papillomavirus genomes amplified in RCA reactions can be linearized by digestion with restriction enzymes (e.g. EcoRV or SacI in the case of CcaPV1 and red deer PV) and used for subsequent cloning and visualization.
In situ demonstration of papillomavirus DNA or antigen in the epithelial layer of skin tumours by i n situ DNA hybridization or immunohistochemistry, respectively, can be applied to investigate the association of papillomaviruses with typical lesions (Figure 16.4). The close antigenic relatedness between the large capsid protein (L1) of BPV1 and other deltapapillomaviruses makes it possible to use commercially available anti-BPV1 L1 antibodies for the detection of cervid papillomaviruses by immunohistochemistry®.
MANAGEMENT, CONTROL AND REGULATIONS
No attempts have been made so far to control fibropapil- lomatosis in wild cervids. Selective culling of animals with visible skin lesions and reduction of host density may be attempted as a control measure, but the potential efficiency of such intervention is dubious.
PUBLIC HEALTH CONCERN
As papillomaviruses are highly host-specific, deer papillomaviruses pose no risk to human health.
SIGNIFICANCE AND IMPLICATIONS FOR ANIMAL HEALTH
P apillomavirus cross-infections between farmed ruminants, including farmed deer, have not been detected. With the possible exception of OvPV1, all deer deltapapillomaviruses seem to be strictly host-species-specific, circulating within a single cervid species.