RUMINANT ALPHAHERPESVIRUS INFECTIONS
CARLOS G. DAS NEVES
Norwegian School of Veterinary Science, Department of Food Safety and Infection Biology, Section of Arctic Veterinary Medicine, Tromso, Norway
The subfamily Alphaherpesvirinae includes several viruses that cause a range of diseases in members of the suborder Ruminantia.
With the exception of Bovine herpesvirus 2 in the genus Simplexvirus, a virus with little impact in European wildlife, all other relevant ruminant alphaherpesviruses are in the genus Varicellovirus, as shown in Table 1.2. Of these, Bovine herpesvirus 1 (BoHV1) is by far the most studied, serving as model for this group of ruminant viruses.BOVINE HERPESVIRUS 1
BoHV1 is the aetiological agent of infectious bovine rhinotracheitis (IBR), and infectious pustular vulvovaginitis (IPV) or infectious pustular balanoposthitis (IPB). BoHV1 causes significant economic losses for the cattle industry worldwide, for which programmes of eradication and/or control of the disease have long been in place.
Although there are several differences in genomic organization and sequences between the different ruminant alphaherpesviruses, mechanisms related to gene expression
TABLE 1.2 Ruminant alphaherpesviruses in the genus Varicellovirus (with permission from Das Neves, 2009(34)).
| Virus | Natural host | Disease | Geographic distribution | Status in European wildlife |
| Bovine herpesvirus 1 | Bovine | Bovine rhinotracheitis, pustular | Europe, America, Asia | Suspected but virus never |
| BoHV1 | (Bos taurus) | vulvovaginitis and balanoposthitis | and Oceania | isolated |
| Bovine herpesvirus 5 | Bovine | Bovine encephalitis | Europe, America, | Not described |
| BoHV5 | (Bos taurus) | Oceania | ||
| Bubaline herpesvirus 1 | Water buffalo | No clinical disease | Europe, Australia | Not described |
| BuHV1 | (Bubalus bubalis) | |||
| Caprine herpesvirus 1 | Goat | Vulvovaginitis, abortion, neonatal | Europe, America, | Suspected but virus never |
| CpHV1 | (Capra aegagrus) | systemic infection, conjunctivitis | Australia | isolated |
| Cervid herpesvirus 1 | Red deer | Ocular syndrome | Europe | Virus isolated |
| CvHV1 | (Cervus elaphus) | |||
| Cervid herpesvirus 2 | Reindeer | Ocular syndrome, respiratory | Europe | Virus isolated |
| CvHV2 | (Rangifer tarandus) | disease, mucosal lesions, abortion | ||
| Elk herpesvirus 1 | Elk | No clinical disease | America | Not described |
| ElkHV1 | (Cervus canadensis) |
or viral replication and latency, as well as pathogenesis, have been shown to be common to all of them.
The BoHV1 genome consists of a double-stranded linear DNA sequence with 135 301 nucleotides, comprising 67 unique genes. Some of these genes encode envelope proteins commonly called glycoproteins. Of these, gB not only plays an essential role in virus attachment and entry, but is also highly immunogenic, representing a dominant viral antigen that can lead to a protective immune response.
Despite its worldwide spread in domestic cattle and being the target of intense study, BoHV1 has not been reported to naturally cause disease in wildlife. Wildlife species have been screened using serological kits for BoHV1 based on gB as antigen. These tests, however, do not enable discrimination between the various ruminant alphaherpesviruses, so it is not possible to rule out the possibility that many wildlife species may harbour herpesviruses closely related to BoHV1 rather than BoHV1 itself.
Seropositive results against BoHV1 have been described throughout Europe in ibex ( Capra ibex), chamois (Rupic- apra rupicapra), red deer ( Cervus elaphus), roe deer ( Capre- olus capreolus), reindeer (Rangifer tarandus), fallow deer (Dama dama), mouflon ( Ovis musimon), European bison (Bison bonasus) and water buffalo (Bubalus bubalis)(35).
Several studies have focused on BoHV1 infections of heterologous hosts. Goats can be infected with BoHV1, leading to high excretion titres and latency. Experimental reactivation has also been successful, but these studies focused on domestic goats and little is known about BoHV1 infections of wild goats, for example. BoHV1 infections of deer and reindeer lead to minimal or no excretion, and latency does not occur. Altogether, studies seem to demonstrate that although BoHV1 can infect some wild ruminants it cannot be maintained over time, and wildlife seem therefore not to be important reservoirs for this virus.
I n primary infections, BoHV1 has potential ports of entry in the nasal cavity, oropharynx, eyes and genital tract.
Replication normally takes place in the epithelial cells, and high titres of BoHV1 are excreted at those ports of entry within 4—5 days post-infection. Although, as for most ruminant alphaherpesviruses, viraemia is possible, it seems that BoHV1 shows little systemic spread and is often restricted to the local ports of entry in primary infections. Nonetheless, BoHV1 can spread by association with mononuclear blood cells and reach the digestive tract, ovaries and fetus, where it can lead to abortion.A strong humoral and cell-mediated response develops within 5 days post-infection, with maximum antibody titres around days 10—12 days post- infection. Residual antibody titres can be detected for up to 2 to 3 years postinfection. Besides the viral lytic cycle, in which active replication takes place, BoHV1 can become latent when viruses migrate to the CNS ganglia (e.g. TG or sacral ganglia) and enter a dormant stage. Different stimuli such as transport stress, calving, and other concurrent infections may lead to reactivation of BoHV1, with the virus returning to the port of entry or spreading to other organs and replicating. Although reactivation can lead to re-excretion, reactivation episodes are often subclinical and hence difficult to identify.
In cattle, IBR can cause clinical signs such as high fever, anorexia, coughing, excessive salivation, nasal discharge, conjunctivitis, dyspnoea and nasal lesions that often consist of clusters of necrotic lesions on the mucosa. In the IPV/IPB form, clinical signs often start with frequent urination and mild mucosal irritation, and can progress to swelling of the vulva with small papules, followed by erosion and ulcers on the mucosal surface of either the vagina or the penis and prepuce. Respiratory infection can result in abortion or neonatal death, especially in calves deprived of colostrum. Infections with BoHV1 alone do not usually cause death in healthy mature cattle, unless in situations where the virus causes a generalized, systemic infection.
No clinical signs or pathology related to BoHV1 infections have been reported in wildlife in Europe.Definitive diagnosis is achieved by viral isolation, detection of viral DNA by specific PCR or restriction endonucleases analysis. Presence of virus in lesions can also be demonstrated by immunohistochemistry or electron microscopy. gB-based serological assays have been designed as an easy tool to identify the presence of BoHV1 infections, especially as antibody titres raised against gB tend to persist for long periods after infection, even though this method will also recognize antibodies against other closely related ruminant alphaherpesviruses. Seroneutralization assays or ELISA with less conserved antigens may be, in these cases, an important additional tool in discriminating among these ruminant alphaherpesviruses.
Controls and treatments for IBR/IPV in cattle vary among European countries depending, among other factors, on the status of the disease in a given country. Options may include the culling of seropositive animals in areas of low seroprevalence, or large- scale vaccination programmes in areas of higher seroprevalence. Identification of latent carriers also constitutes an important step in the control and prevention of outbreaks of BoHV1.
There are no known public health concerns associated with BoHV1. The impact of cross-infections between ruminant alphaherpesviruses is described in the sections on caprine and cervid herpesviruses.
CAPRINE HERPESVIRUS 1
Caprine herpesvirus 1 (CpHV1), previously known as Bovine herpesvirus 6, is an alphaherpesvirus closely related to BoHV1. It is widely distributed across Europe, can cause enteritis and generalized infections in neonatal kids, and induces vulvovaginitis, balanoposthitis, respiratory disease or abortion in adult animals.
This virus should not be confused with Caprine herpesvirus 2 (CpHV2), a recently classified gammaherpesvirus closely related to Ovine herpesvirus 2 and Alcelaphine herpesvirus 1, which is endemic in domestic goats and has been observed or is suspected to cause clinical MCF in certain species of deer in the USA(36) and Norway(26).
CpHV1 has a genomic organization similar to the genome of BoHV1, and there are several different strains.CpHV1 probably has a worldwide distribution. In Europe, studies have reported this virus, or antibodies against it, in domestic goats in Italy, Spain, Norway, Greece, Germany and France (Corsica). CpHV1 is genetically and antigenically closely related to BoHV1, and conventional ELISA testing for antibodies usually cannot differentiate the virus to which the immune response has developed.
Few studies have been carried out in wild populations, but in France ibex ( Capra ibex), chamois (Rupicapra rupi- capra), red deer (Cervus elaphus), and roe deer (Capreolus capreolus) have tested seropositive for CpHV1(37).
Pathogenesis of CpHV1 is very similar to that of other ruminant alphaherpesviruses. CpHV1 can infect the animal by either the respiratory or the genital routes, quickly establishing viraemia and being detected in a variety of organs. Latency takes place at the TG or sacral ganglia, and upon reactivation (e.g. physiological stress), respiratory, genital tract and ocular re-excretion of the virus is possible. Experimental reactivation of CpHV1 has only been successful under high doses of dexamethasone. In kids the virus spreads very quickly, leading to a systemic disease with high morbidity. Vertical transmission from mother to fetus is also possible, and CpHV1 is associated with episodes of abortion in domestic goats, especially during the second half of pregnancy(38). The impact of CpHV1-related abortions in wild goats and other members of the Caprinae subfamily remains unknown.
CpHV1 can lead to four main types of clinical situations.
1. Systemic form, often seen in young kids that exhibit progressive weakness and abdominal pain. There can be conjunctivitis and purulent nasal discharges, erosion in the oral and nasal cavities and oedema of the myocardium, among other signs. Animals usually die within 2—4 days after the onset of the infection1-39).
2. Genital form, mostly in adult animals, where the infection can often be subclinical. Lesions in the genital mucosa can develop with hyperaemia, oedema and the appearance of papules, vesicles and ulcers(40).
3. Respiratory form, usually combined with a secondary bacterial infection, where animals develop an acute pneumonia(41).
4. Abortive form, especially during the second half of pregnancy, frequently without any other clinical ill- health in the dam, or lesions in the aborted fetus(38).
There is no known treatment for caprine herpesvirus infections. Confirmation of CpHV1 infection is usually achieved by viral isolation, or detection of viral DNA by specific PCR. Presence of virus in lesions can also be demonstrated by immunohistochemistry or electron microscopy. Conventional ELISA screening serology for BoHV1 will recognize antibodies against CpHV1 but not distinguish between the two viruses. Seroneutralization assays may be, in these cases, an important additional tool.
Given the potential of CpHV1 to cause fatal diseases in kids and also in adult animals, hygiene-based preventative control measures in the event of an outbreak are essential to avoid the rapid spread of the virus. The culling of seropositive animals and latent carriers may be necessary to prevent spread of disease. Vaccination studies are underway and, as for BoHV1, this might prove to be an important tool for the control of outbreaks.
There are no known public health concerns associated with this virus. CpHV1 has been shown to infect and establish latency in bovine calves, but reactivation was not successful and clinical signs were absent(42). The impact of this virus in other species of the Caprinae subfamily remains unknown.
CERVID HERPESVIRUSES 1 AND 2
Serological studies of cervids have long shown exposure of these animals to BoHV-1 or other closely related viruses. Two ruminant herpesviruses belonging to the same Alphaherpesvirinae subfamily as BoHVl have so far been identified and isolated in Europe from members of the Cervidae family: Cervid herpesvirus 1 (CvHVl) in red deer (Cervus elaphus) and Cervid herpesvirus 2 (CvHV2) in reindeer (Rangifer tarandus).
CvHV1 and CvHV2 can cause outbreaks of infectious keratoconjunctivitis in red deer and reindeer, respectively(43). CvHV2 has further been demonstrated to have the potential to be involved in respiratory disease and abortion(34,44).
Two different strains of CvHV1 have so far been identified in the UK and Belgium1-45’46), while isolates from CvHV2 in Norway and Finland seem to represent the same strain1-47’48). Both viruses have a genomic organization similar to that of BoHV1(49).
Because of the close genetic and antigenic relationship between cervid herpesviruses and BoHV-1, serological cross-reactions are detected by conventional ELISA testing. Because of this, many surveys that have classified cervids as seropositive for BoHVl may have actually detected antibodies against cervid herpesvirus. CvHV1 and CvHV2 may hence be much more common in cervid populations than previously thought.
CvHV1
Cervid herpesvirus was first identified during an outbreak of ocular disease in farmed red deer in the UK in 1982(45), but since then many serosurveys have identified the presence of alphaherpesviruses circulating among different deer species. Studies revealed high seroprevalences among red deer in England as well as in the Czech Republic (translocated animals tested by virus neutralization in the early 1990s). A serosurvey of wild animals in France and Belgium revealed higher neutralizing titres against CvHV1 than BoHV1(37). Other studies have also identified seropositive red deer in Scotland, England, France, Norway and Germany. Some roe deer (Capreolus capreolus) were also found to be seropositive for either CvHV1 or a BoHV1- related virus in France, Germany, Norway and Hungary, where fallow deer (Dama dama) were also found to be seropositive.
CvHV2
Serosurveys in semi-domesticated reindeer (Rangifer tarandus tarandus) in Finland, Sweden, Norway and Greenland have all revealed the presence of a BoHV1- related virus. Seroprevalences have ranged from 12% in Finland in 1977(50) to more than 48% in Norway in 2003-2006(52). Although most reindeer in Scandinavia are semidomesticated, meaning that although free-ranging they are owned and herded, serosurveys on wild reindeer populations in both Greenland and Norway have also shown these animals to be infected1-52’53). Recent Seroneutralization tests in Norway determined much higher titres against CvHV2 than BoHV1, a fact also confirmed by CvHV2 isolation in 2009. The same studies have shown that age and animal density represent risk factors for CvHV2 infections)51).
The reindeer subspecies R. tarandusplatyrhynchus inhabiting the Svalbard archipelago in the high Arctic was screened in the early 1990s’ with all tested animals being seronegative)54).
Like other alphaherpesviruses, CvHVl and CvHV2 are transmitted directly by close contact between infected animals’ e.g. licking’ nuzzling’ sneezing or venereal contact. Both viruses have the potential to infect the genital and respiratory tracts as well as the ocular mucosa’ as demonstrated either by disease outbreaks or experimental studies of CvHVl in England and France, and CvHV2 in Norway and Finland)45’47’55-58). In experimental infections with either CvHVl or CvHV2, animals usually display a transient hyperaemia in the mucosae during the initial period of viral excretion’ with increased nasal and genital discharges in some cases)55,56,59).
F or both agents’ viral excretion titres reach their maximum between 4 and 7 days post- infection at the point of entry )respiratory or genital tract), decreasing quickly after that as the humoral response commences at around 10 days post-infection.
Although little is known about the viraemic potential of CvHV1, in the case of CvHV2 experimental studies show that after genital or respiratory infection the virus spreads throughout the body, reaching different organs, such as liver, lung, spleen and lymph nodes)44,56,57). Vertical transmission was observed in several animals.
Both viruses have been shown experimentally to become latent in their hosts and reactivation, excretion and reisolation has been observed experimentally)34,55,57).
Reindeer infected with CvHV2 can exhibit lesions in the skin of the eyelids, lips and gingiva, as described in Norway)48). In the same experimental study in Norway, one abortion occurred, with CvHV2 being recovered from both maternal and fetal tissues. This strengthened the hypothesis that CvHV2 contributes to, or causes, abortion. CvHV2, like BoHV1, seems to have the potential to be involved in respiratory disease complex, where reactivation of the virus can lead to it reaching the respiratory pathways, with immunohistological findings showing it to be associated with epithelial hyperplasia and destruction)56).
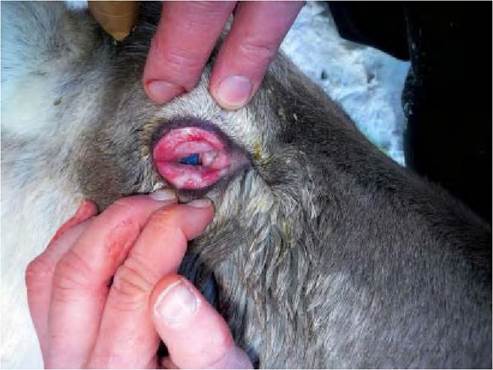
FIGURE 1.4 Infectious keratoconjunctivitis caused by Cervid herpesvirus 2 infection in semi-domesticated reindeer during an outbreak in Norway in 2009. Severe degree of periorbital oedema. With permission from Das Neves, C.G. et al., 2010)48).
Outbreaks of keratoconjunctivitis have been reported to be associated with both CvHVl and CvHV2 in red deer and reindeer in Scotland )1982) and Norway )2009), respectively)45,58). In Scandinavia, similar outbreaks of ocular disease in reindeer have been recorded for more than l00 years.
Ocular disease is characterized, both in deer and reindeer, by purulent ocular discharge, hypopyon, uniform corneal opacity without ulceration, mucopurulent nasal discharge and photophobia. Moderate swelling of the periorbital tissues and marked oedema of the eyelids are also observed )Figure 1.4). In more severe cases, there is a secondary bacterial infection and haemorrhagic and purulent exudates appear, leading, in extreme cases, to blindness and destruction of the eye. Moraxella bovis has been reported in this ocular syndrome in reindeer in Finland, but in an outbreak in Norway in 2009 Moraxella boviculi was reported for the first time in reindeer. Because of the potential of many ruminant alphaherpesviruses to crossreact serologically, only tests based on virus detection such as isolation, viral DNA detection or restriction endonuclease analysis can accurately determine which virus is present in a given animal or population. The presence of viral particles in lesions can also be demonstrated by immunohistochemistry or electron microscopy.
There is no known treatment for cervid herpesvirus infections. When clinical signs are present, infected animals should be isolated, as the virus spreads easily horizontally.
There are no known public health concerns associated with Cervid herpesvirus 1 or 2.
Cervid herpesviruses seem to pose little risk for domestic ruminant populations. Experimental studies indicate that bovines seem to be refractory to CvHV1 infection but can be infected with CvHV2, which can cause a mild rhinitis without latency or reactivation(59). Both crossinfections have never been reported outside experimental conditions. In southern Scandinavia, reindeer and deer share common grazing areas; however, cross infections between CvHV1 and CvHV2 in these two species has not been studied.