Specific immunity
Despite the impressive benefits of nonspecific defenses, this alone is not sufficient. Hallmarks of both cellular and humoral immunity include (1) specificity, (2) systemic rather than local responses, and (3) evidence of memory.
Specificity indicates that the immune response is directed at a unique antigen. Systemic responsiveness refers to the notion that a response to an attack can be mounted regardless of the point of entry. Memory indicates that the immune system is better prepared with a faster response when exposure to an antigen occurs a second time. This is the idea behind vaccinations. A vaccination is essentially the induction of a specific immune response and creation of immunological memory as a result of a planned exposure to the antigen in a manner that does not cause illness. For example, immunization with cell wall components or killed bacterial cells (incapable of causing disease) can nonetheless induce an immune response because of the foreign proteins or polysaccharides present in these preparations. If the animal is then exposed to live microorganisms, the animal can respond more quickly so that the chance of exhibiting the disease is reduced or symptoms are milder and less severe compared with nonimmunized animals.Let's first consider humoral or antibody-mediated immunity. However, it is worth remembering that the two divisions of the immune system (humoral and cell-mediated) do not function independently but rather function together to enhance protection. As Figure 15.8 shows, it is also important to realize that immunity can be either passive or active. Clearly, the protection provided to the newborn calf (or puppy in our earlier example) by suckling colostrum from its
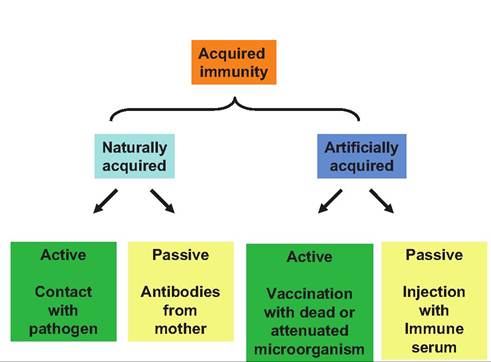
Fig. 15.8. Types Ofacquirecl immunity. Both naturally and artificially acquired immunity can be very important, but only active immunity can establish immunological memory.
Once antibodies from the mother or via injection are degraded, they cannot be replaced by the animal.Box 15.2 Natural killers
Just the idea of some of our animals' lymphocytes "running around" with the capacity of kill other cells seems somehow unnerving. The notion that selected T cells can be induced to target virus- infected cells or tumor cells is more comforting. As reviewed by Boysen and Storset (2009), so-called natural killer (NK) cells were first discovered in the 1970s as cells that could spontaneously lyse and kill tumor cells without prior immunization. It was subsequently recognized that the cells that attack cells cannot be "identified" as self (i.e., the absence of surface markers common to somatic cells of an individual). Essentially, if the NK cells do not recognize a self-MHC class I molecule on a target cell, no inhibitory signal is delivered, and the target cell is doomed.
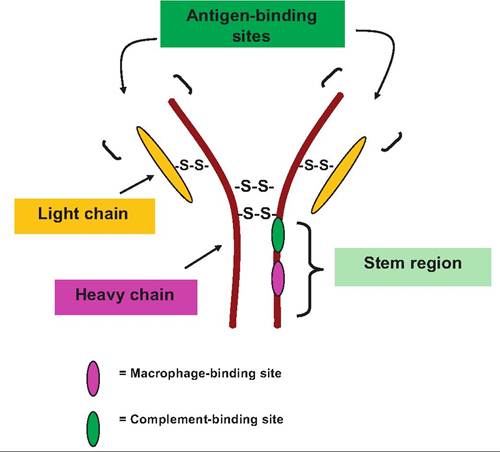
Fig. 15.9. Basic antibody structure. An antibody molecule consists of four protein chains: two heavy and two light chains. The portion of the heavy chain resembling the stem of the letter Y provides sites of activation of complement and interactions with macrophages. The combination of light and heavy chains at the ends of the arms of the Y creates two identical antigen-binding sites. Disulfide bonds are prominent in linking heavy and light chains together as well as at locations within the chains.
mother is critical, but it is limited. The antibodies provided in the colostrum are only a stop gap measure until the calf or puppy is able to begin mounting its own immune responses with related immunological memory (Box 15.2).
Antibody structure and function
There are five major classes of immunoglobulins: IgA, IgG, IgM, IgD, and IgE. IgG is the most abundant and diverse antibody in circulation. It accounts for -80% of the total. It is primarily responsible for both primary and secondary antibody responses (increased blood titers) following immunization or other exposure to antigens.
IgG also circulates as a monomer. IgA also appears as a monomer but is limited in the circulation. It occurs more frequently in secretions (saliva, sweat, intestine, etc.) associated with mucous membranes and epithelial surfaces. It also is most frequently secreted as a dimer with the two antibody molecules joined by a third element called the secretory piece. IgM appears as a monomer and in groupings of five antibodies (pentamer) linked together. In the monomeric form, the antibody is usually attached to the surface of B cells. During primary antibody responses, the pentamer form of IgM is the first class of antibody released by the plasma cells. Since there is usually only a small amount of IgM free in the circulation, detection of an increase in the blood is diagnostically useful as an indicator of a current infection in an animal. IgD is nearly always attached to the surface of B cells where it functions as a receptor for the activation of the cells. IgE is secreted by plasma cells in the skin and mucosal membranes. When the IgE is bound to antigen, the stem region of the molecule reacts with mast cells and basophiles, causing them to release histamine and other chemicals. Unfortunately, hyper or inappropriate IgE-induced activation of these cells is involved in many allergic reactions. This explains the significance of antihistamine treatments to treat allergic reactions.Whatever its specific class, each antibody molecule has a basic structure consisting of four protein chains linked together by disulfide bonds. Two identical heavy chains of about 400 amino acids each make up the bulk of a structure that resembles the shape of the letter Y. Two additional identical protein chains (the light chains) essentially overlap the portions of the heavy chains in the arms of the Y and are joined by sulfide bonds. The heavy chains have a hinge-like region near the top of the stem of the Y The two ends at the top of the Y of the antibody molecule create the sites for binding of the antibody to its antigen.
This means that the antibody is divalent; that is, one antibody molecule is capable of binding two antigen molecules. The stem region of the molecule is significant because it contains sites for complement binding and for macrophage activation. These sites are important because the binding or fixing of the antibody allows the development of a cascade of reactions important in the immune attack. The structure of an antibody molecule is illustrated in Figure 15.9.Although antibodies do not directly destroy pathogens, their binding to antigens on the surface of bacterial cells, for example, marks the cells for destruction. Antibody binding to toxins or foreign debris can also inactivate these agents by neutralization, precipitation, or, in the case of cell-associated antigens, agglutination. These reactions greatly enhance inflammation, and along with inflammation, induced chemotaxis recruits leukocytes that destroy bacterial cells by phagocytosis. Antibody binding also triggers complement fixation and exposes the macrophage-binding sites on the antibody molecule. The coating of foreign substances with antibodies is called opsonization.
Antigens are substances that can activate the immune system, thereby provoking an immune response. Most often these are large, complex molecules (typically proteins) that do not normally appear in the body. In other words, the immune system does not recognize them as self. In the case of completely reactive or immunogenic antigens, these molecules induce proliferation of specific lymphocytes and the synthesis and secretion of specific antibodies. In other cases, many small peptides, nucleotides, are not immunogenic in themselves, but when linked with other self-proteins, the new combinations can produce a dramatic, even harmful, response. This is the basis of some allergenic responses. Researchers were able to take advantage of this property to create antibodies against normally nonresponsive molecules, that is, steroid hormones to create many immunological-based assays, for example, radioimmunoassay, enzyme-linked immunoassays (ELISA), and western blotting.
B cell selection and antibody secretion
When B cells are stimulated by antigens, a cascade is initiated so that antigen binding to surface receptors on a particularly naive or previously inactivated B cell becomes activated to complete its differentiation cycle. This activation process, usually in combination with T cells, triggers clonal selection and expansion. The initial step induces B cell growth followed by rapid proliferation of an army or clone of identical daughter cells, all of which express receptors specific for the antigen that initiated the process. Since all of the cells are identical, they form a group called a clone. In cases where a particular antigen leads to the production of a single clone of cells, this would be called a monoclonal response. This situation is taken advantage of experimentally to produce highly specific monoclonal antibodies. More often in physiological situations, several different families or clones of stimulated B cells are generated. Since each of the antibodies that are produced is likely to recognize different epitopes
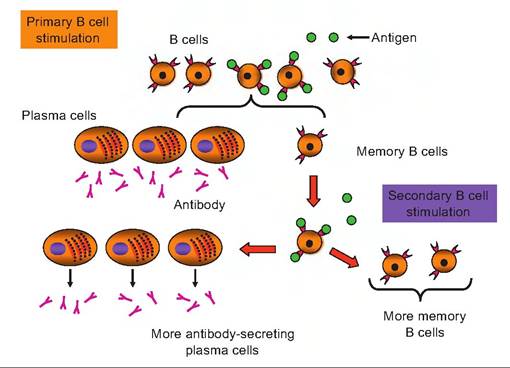
Fig. 15.10. Antigen stimulation of B cells. In the primary stimulation, antigen (green spheres) binds to receptors on the surface of selected B cells. This induces cell proliferation and production of clones of identical cells. Some of the cells enlarge into B lymphoblasts and then plasma cells that secrete antibodies specific to the antigen. Other clonal B cells remain as memory B cells. With a subsequent exposure to the antigen (weeks, months, or even years later), a second, more rapid induction and secretion of antibody and generation of additional memory B cells occurs.
of the same antigen, these antibodies are referred to as polyclonal antibodies.
Most of the stimulated B cells are induced to become plasma cells. This is fortuitous because plasma cells have an extensive array of rough endoplasmic reticulum (RER).
Thus, the capacity of these plasma cells to synthesize and secrete antibody molecules (proteins) is very high. This is illustrated in Figure 15.10.It is estimated that each plasma cells can secrete more than 2000 antibody molecules per second. These antibodies have the same antigen-binding capacity as the receptor proteins on the surface of the B cells that first bound the antigen. Clonal B cells that are not activated to become plasma cells become long-lived memory cells. It is the presence of the memory B cells that explains the very rapid and sustained secretion of antibody that occurs when an animal is exposed to an antigen for a second time. This pattern of response is illustrated in Figure 15.11.
Cell-mediated immunity
Antibodies are extremely important, but it is also clear that effectiveness depends on the capacity of the antibodies to recognize specific pathogens. How are our animals protected against viruses or infections from microorganisms that can "hide" from detection by antibodies? The T cells provide this added, more complex layer of protection. Two primary groups of T cells have been recognized based on the complexes of
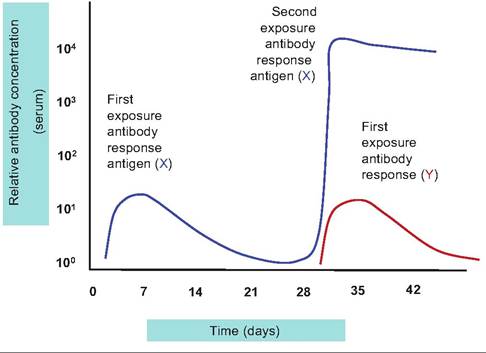
Fig. 15.11. Primary and secondary antibody responses. The blue line illustrates the relative antibody secretion response to immunization with antigen X. The initial response begins after several days, peaks at a relatively low level, and declines markedly by 4 weeks. A second exposure to antigen X at this time elicits a rapid and relatively much greater response. Exposure to a second antigen, Y, at the same time has no effect on the response to antigen X, and the relative response to antigen Y is similar to the initial reaction noted to immunization with antigen X.
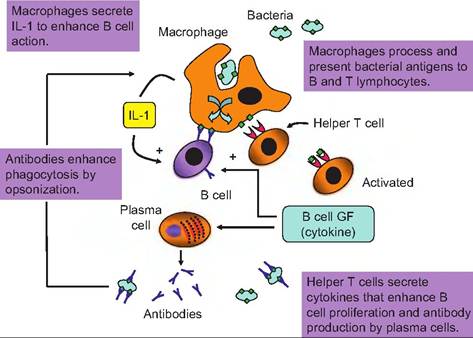
Fig. 15.12. Antigen process in macrophages. Bacterial cells are engulfed by macrophages, and antigens are processed and expressed on the surface of the macrophage. Helper T cells associate with the macrophage and bind fragments of bacterial cell proteins "presented" by the macrophage. This activates the helper T cell so that it is stimulated to secrete cytokines which stimulate proliferation of B cells and conversion into plasma cells that secrete antibodies specific to the bacterial antigen.
glycoproteins that are expressed on their cell surfaces. These are CD4 cells, also known as helper T cells (Th) and a larger population the CD8 cells that are mostly cytotoxic T cells (Tc). Some of the CD8 expressing T cells are also suppressor (Ts) cells that act as modulators of cell-mediated immunity.
While B cells and antibodies bind to and respond directly to antigens, T cells do not have this ability. Instead, the T cells can recognize and respond to pieces or fragments of protein antigens that have been processed by other cells of the immune system, the antigen-presenting cells (macrophages, neutrophils, dendritic cells of the skin, etc.). This process is illustrated in Figure 15.12.
Cytotoxic T cells (Tc), also called killer T cells, have the ability to directly attack and kill other cells. When activated, these cells migrate through the circulation and the lymphoid tissues seeking cells that express antigens for which the Tc cells have been sensitized. The primary targets of the cells are other cells that have been infected by viruses, but under some circumstances they can attack cells that are infected by bacteria or parasites. They can also act on cancer cells and are the primary cells involved in transplant rejection reactions.
Before the cells can respond, the Tc cells have to link or dock with potential targets by binding to the self/ non-self-complex on the cell surface. Briefly, the surface of all cells expresses a myriad of proteins. However, if the immune system has been appropriately programmed, all of these self-antigens are not recognized as foreign by the particular animal but are strongly antigenic to other animals. This is the essence of blood transfusion or graft rejection between unrelated animals. Some of the major surface proteins are part of a group of glycoproteins called major histocompatibility complex or MCH proteins that are coded by MHC genes. Because there are virtually millions of possible combinations for the complex of genes that code for these proteins, except for identical twins, it is very unlikely that two animals would have expression patterns that would be the same between individual animals. It is even more complex in that there are two major clusters of MCH proteins and genes. Class I MHC proteins are expressed on surfaces of essentially all cells but class II MHC proteins appear only on certain cells of the immune system. Each MHC protein has a cleft or groove that displays a peptide. In normal cells, the peptides that are displayed in this cleft are peptides that are derived from normal recycling of cellular proteins. When some cells become infected or are altered by cancer, the MHC proteins can bind and display peptides derived from bacterial cells, viruses, or cancer-mediated processes. This then acts to mark the infected or cancerous cells as nonself, thereby targeting them for close surveillance and possible attack by the Tc cells.
Essentially, the CD8 class of lymphocytes or Tc cells is activated when they dock with other cells that have processed antigens in combination with the class I MHC proteins that are not recognized as self. The example shown in Figure 15.13 illustrates the activation of Tc cells that have encountered a virus-infected
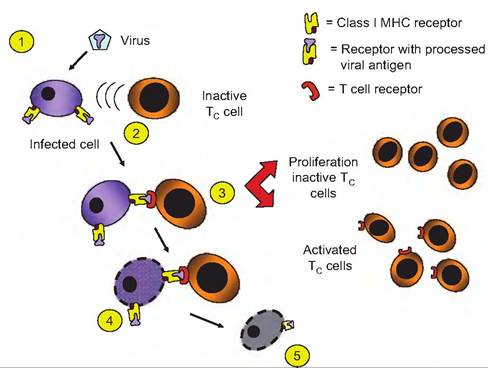
Fig. 15.13. Cytotoxic and helper T cells activation. Both Tc and Th cells are stimulated to proliferate and produce clones when they associated with foreign antigens (1) that are in complexes with MHC proteins on cell surfaces. In this example, a competent Tc cell docks with a cell expressing a foreign viral antigen in combination with the cells MHC proteins (2). Activation (3) induces proliferation leading formation of more Tc cells (memory) as well as currently activated cells. When the activated Tc cells complexes with an infected cell (4), the infected cell is ultimately (5) destroyed. Activation of Th cells would occur in a similar fashion except the complex would involve class Il MHC proteins rather than class I MHC proteins.
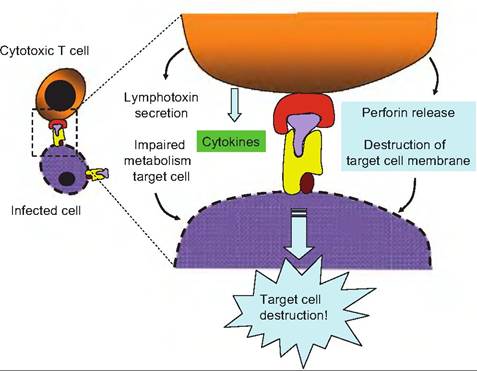
Fig. 15.14. Cytotoxic T cell attack. Once the Tc cell has encountered and responded to an infected cell, several different mechanisms can be induced to produce destruction of the target cell. Release of Iymphotoxin can essentially poison the cell by disrupting its metabolism. Release of perforin can act to perforate and disrupt the plasma membrane of the cells. Granzymes released by T cell vesicles (granules) can degrade target cell proteins. Finally, some cytokines released act to promote apoptosis (programmed cell death in the target cell).
cell and, as a consequence, a clone of activated Tc cells is produced. These cells can then detect and bind to other infected cells. The fundamental result is that this encounter induces the Tc cell to divide and produce clones of identical cells. Some of these new cytotoxic T cells become memory T cells, while others are activated to seek out cells that exhibit the antigen/MHC on the cell surface that initiated the process in the beginning.
The actual mechanism for the Tc destruction of the "foreign" cell is complex and poorly understood, but several events occur at least in some cases. When binding between the cells occurs, the Tc cell releases granules that contain a protein called perforin that inserts itself into the plasma membrane of the cell targeted for destruction. Especially in the presence of Ca++, the perforin molecules combine to create pores through the plasma membrane. This allows entrance of inactivated enzymes called granzymes, also contained in the vesicles released from the T cell, to enter the cytoplasm of the targeted cell. Once inside the target cell, like a miniature Trojan horse, the proteases are activated, and the cellular machinery begins to degrade. In other cases, the Tc cells can produce the death of targeted cells by secretion of Iymphotoxin, a protein that causes the fragmentation of the target cell's DNA. Other Tc cells release γ interferon which activates macrophages in the area to killer status so that the death of the targeted cells is indirect. This is analogous to hiring a hit man to do the deed. Details of Tc attack are illustrated in Figure 15.14.
A final type of T cell to consider is the suppressor (Ts) cell. As the name suggests, these cells act to dampen the aggressive action of the Tc cells once the inflammation has subsided. In addition, it is believed that the Ts cells are important in minimizing autoimmune reactions, that is, in situations where the immune system can attack normal healthy cells. However, suppression does not occur immediately. In part this is because most of the activation that initiates the T cells to respond in the beginning increases the number of Tc and Th cells to a much greater extent than it does for Ts cells. Table 15.2 summarizes the immune cells that participate in defense of tissues and cells.
Avian versus mammalian systems
At a global level, many aspects of immunology are similar between species, for example, the presence of humoral versus cellular immunity, significance of circulating antibodies, impact of inflammation, and the role of cell signaling in maturation of various classes of leucocytes. However, understanding of immune responses in avian species, especially chickens, is especially significant scientifically. In 1956, Glick et al. reported that the chicken bursa of Fabricius (BF) was essential for development of humoral immunity.
Table 15.2. Immune cells involved in defense of tissue and cells.

Bursectomized chickens had no antibody production responses when challenged.
The BF is an invagination of a portion of the cloaca. It contains 10-15 folds surrounding a lumen with tissue containing approximately 10,000 follicular structures. These follicles develop in part from stem cells that migrate from the yolk sac and embryonic liver to the BE As development progresses, BF structures change from small buds to colonies of clonal B cells that proliferate and move to the outer regions of the BE Complex gene rearrangements and gene conversion events ultimately create populations of potential B cells. Complicated signals associated with cytokines specific to the bursa and cell interactions leave only about 5% of these cells to become fully functional B cells that are active in humoral immunity (Scott, 2004). Thus, the identification of these cells as B lymphocytes reflects the fact that in birds, these cells develop in the BE It also distinguishes these lymphocytes from T cells which are thymus derived. Mammals do not have a structure equivalent to the BE Consequently, the bone marrow serves as a site for B cell maturation.
As you might guess, B cell development occurs in progressive steps, but based primarily on mouse models, each stage depends on dramatic gene alterations. As discussed earlier, the antibody molecule is composed of two light (L) and two heavy (H) chains. Moreover, the genes directing the synthesis of these proteins are found in corresponding L and H chain loci. An H chain locus has three regions, V, D and J, which recombine randomly as the cells mature. This creates unique variable domains in the immunoglobulin that can be made by each individual B cell. Similar rearrangements occur for L chain locus, but there are just two regions [V and J]. The outline given below briefly describes events during sequential developmental stages of B cell maturation:
• Progenitor B cells have starting germ cell H and L genes.
• Early Pro-B cells get beginning of D and J rearrangement on the H chains.
• Late Pro-B cells have beginning of V-DJ rearrangement of V, D, J on the H chains.
• Large Pre-B cells occur as the H chain is VDJ rearranged, germ line L genes.
• Small Pre-B cells undergo V-J rearrangement on the L chains.
• Immature B cells have the VJ rearranged on L chains, VDJ rearranged on H chains, and expression of IgM receptors occurs.
• Mature B cells begin expression of IgD.
If the B cell fails in any step of the maturation process, apoptosis leads to the death of the cell. Moreover, if the cell recognizes a self-antigen during the maturation process, the cell either becomes arrested or undergoes apoptosis. This probably explains why only a small percentage of precursor B cells survive to become part of the peripheral B cell pool.