Innate or nonspecific defenses
As the name suggests, nonspecific defenses do not distinguish one threat from another. The protection that is afforded is the same no matter what the circumstances. These measures include physical barriers, phagocytic cells, complement, general inflammatory response, and fever.
The first line of defense against pathogens at first blush seems minor, but in reality, the protective properties of the skin and the mucous membranes are substantial. Consider the physical barrier that intact skin provides and the problems that rapidly occur if there are cuts or open wounds. The stratified nature of the skin epithelium, presence of desmosomes to link the cells together, keratinized surface, and secretions of sebaceous and sweat glands are highly protective. The importance of lanolin on the wool of sheep to provide protection from the elements is another example. Extend this to many animals, and the added physical protection provided by fur, hair, or wool becomes apparent. The acidity of skin secretions (pH 3 to 5) also inhibits bacterial growth. Move to the internal passageways of the respiratory, digestive, and urinary tracts. As you should recall, the more exterior portions of these tracts (oral cavity, esophagus, rectum, etc.) are also covered by stratified squamous epithelial cells. In more internal regions, the epithelium is typically thinner and simple, but the cells are linked by tight junctions that increase the barrier function of the epithelium. In addition, specialized glands (globlet cells and various multicellular glands) provide mucus and specific proteins that act to coat and protect the internal surface. Saliva and lacrimal fluids of the eye contain the protein lysozyme and enzymes that can attack bacteria. In mammary secretions produced during the nonlactating or dry period, accumulation of the protein lactoferrin binds iron, which acts to impair growth of bacterial cells. The stomach mucosa, with secretion of HCL and peptidases, also kills many ingested microorganisms.As a specific agricultural example, consider the importance of the epithelial layer of the teat end of the dairy cow in protection of the internal mammary gland from mastitis (inflammation of the mammary gland caused by invading microorganisms). The teat of the ruminant has a single opening called the streak canal that leads directly into a space within the teat called the teat cistern (see Chapter 18). This means that the structure of the streak canal is critical as the primary defensive barrier against mastitis. The lowest 2 cm of the streak canal is especially important because of the capacity of tissues in this region to act as a barrier to minimize milk leakage or entrance of environmental agents. Intuitively, the diameter of the streak canal is positively related to the rate of milk flow, but cows with the best balance of acceptable rates of milk flow and protection from bacterial invasion have the greatest longevity in the herd. Because the teat canal is lined with longitudinal folds, dilation of the streak canal during milking causes the epithelial lining to become flattened and thin during milking. This is analogous to the changes in transitional epithelium in the bladder. Regardless, this periodic stretch allows the keratin to spread over the surface to form a bactericidal barrier. With milking, some of the keratin is flushed away during the periodic opening and closing of the teat canal, but fortunately it is constantly being renewed by the epithelium (Capuco et al., 1990). The keratin itself has antibacterial agents that inhibit the growth of pathogens. Some researchers also suggest that the minute areas of secretory tissue in the area of FurstenbergzS rosette (near the opening of the teat cistern) secrete protein(s) with bactericidal effects, but others suggest the material is Iipid-Iike and made by the epithelial cells secreting the keratin.
Certainly, the epithelial cells of the streak canal are constantly being renewed based on the appearance of mitotic cells in the basal layers of the epithelium (stratum germinativum). Passage of cannula through the teat canal or use of teat dilators scrapes away the keratin and can traumatize the epithelium. Experimentally, resistance to mastitis is markedly reduced if the keratin layer is removed. Studies in which pathogens were inoculated 3mm into the streak canal caused infections in about one-third of treated glands. Inoculations 4mm into the streak canal increased infection rates further, and inoculations 5mm into the streak canal nearly always caused infection. This confirms the significance of this barrier function (Nickerson and Pankey, 1983). Since pathogens, which cause mastitis, are not motile, to gain entrance into the parenchymal tissue they must be moved by physical forces from the outside of the teat, through the streak canal, teat and gland cistern, and larger ducts, and to the alveoli. Other than the period around milking, the keratin of the streak canal makes an effective barrier. However, animals with inherently thin keratin or animals with damaged teat areas are susceptible to local colonization with microbes and are therefore at greater risk to infection. During milking itself, retrograde movement of milk due to vacuum fluctuations or vacuum slips with leakage of air around the teat cups can allow bacteria-laden droplets to pass the streak canal.During machine milking, there are certainly dramatic effects on the teat and teat end. Given the rate of milk flow, 7-8m∕s, it is reasonable to expect that resulting shear forces might remove some of the protective keratin. It is also probable that some milk constituents are absorbed into the keratin during the time of milking or from milk droplets remaining after milking. If milking removes substantial amounts of the keratin and if renewal is delayed or changes in composition favor the formation of bacterial colonies or adherence, this could have marked effects on the streak canal as the primary defense against mastitis.
Phagocytes are cells that engulf pathogens and cell debris. These include neutrophils and eosinophils that normally circulate in the blood and macrophages that typically reside within tissues. Some of these macrophages are considered fixed because they are permanent residents of a tissue. Examples include the Kupffer cells of the liver or the star-shaped dendritic cells of the skin. Other "free" macrophages are more mobile and can respond to problems throughout the body or act to patrol local tissue areas. An example of the latter is the alveolar macrophages of the lung, which patrol the internal surface of the alveoli. Monocytes in the bloodstream are converted into macrophages when they exit the circulation.
Macrophages (monocytes) and microphages (neutrophils, eosinophils) share the ability to migrate and squeeze between the endothelial cells of capillaries by a process called diapedesis. This process is typically initiated by injury to the endothelial cells and/or appearance of factors that act as attractants for the cells. This is called chemotaxis. The process begins with adhesion followed by the leukocytes forming a sort of pavement of cells lined up along the periphery of the capillary. With increased permeability or damage, there is then an often rapid diapedesis of the cells between the endothelial cells to the region where the bacterial cells or there toxins are located. Figure 15.2 illustrates general events that occur with an inflammatory response. Many of these effects are mediated by the release of chemical compounds produced by injured cells, proteins that appear in exudates from the circulation, as well as molecules released by stimulated platelets and phagocytes. The local response to infection is called inflammation but depending on the particular circumstances, the physiological response can become more widespread. Fever, a higher-than-normal body temperature, occurs in response to some microorganisms and their toxins. For example, in dairy cattle, mastitis caused by Staphylococcus aureus species is typically localized, but mastitis caused by Escherichia coli is characteristically associated with marked systemic effects well beyond appearance of abnormal milk and heat and redness of the udder.
Fever, lethargy, absence of appetite, and markedly reduced milk production are common. Neither is it uncommon for systemic problems to become so severe that physiological systems can fail and death occurs. With marked activation of leukocytes exposed to especially potent pathogens, pyrogens secreted by the leukocytes and toxins from the microorganisms act to reset the hypothalamic neurons responsible for homeostatic control of body temperature. Prolonged high fevers are dangerous because excess heat denatures enzymes. However, milder fever is beneficial because metabolic rates of tissues are increased, allowing for more rapid repair and healing. Interestingly, while fever is known to occur in virtually all mammals, responses in birds are poorly understood (Gray et al., 2012).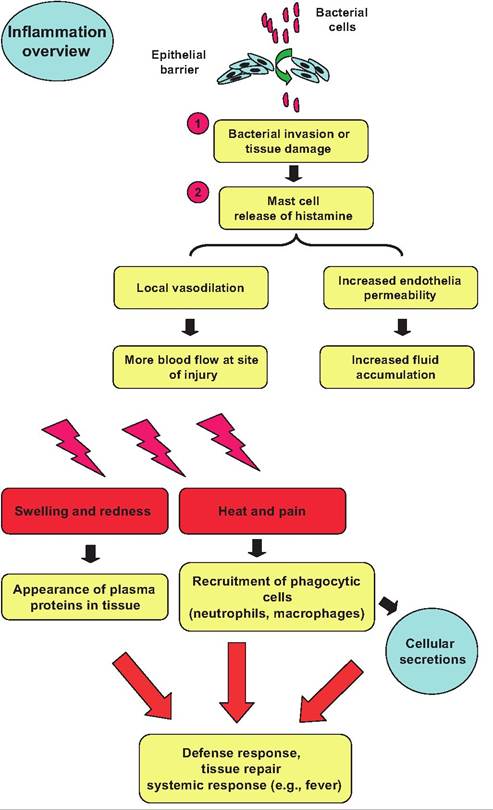
Fig. 15.2. Bacterial attack and inflammation response. Following penetration of the epithelial barrier by bacteria damage to tissues or release of bacterial products, the liberation of histamine, complement, and other molecules initiates a cascade of responses. Increased vasodilation, capillary permeability, and appearance of substances chemotactic to phagocytes produce rapid migration of these cells to the region. The four primary signs of acute inflammation are heat, redness, pain, and swelling.
When infection is severe, responses can be dramatic. The creamy or yellowish pus that fills infected tissue areas is a mixture of dead cells, tissue debris, as well as dead or dying microorganisms. If repair mechanisms do not clear the area of debris, these materials can become walled off by layers of collagen fibers and other extracellular matrix proteins, essentially scar tissue. This is a protective mechanism (e.g., isolation of the affected area), but this can also lead to creation of an abscess. In these cases, it is often necessary to surgically drain the material before healing can take place.
In some particularly difficult infections, bacterial cells can be engulfed by macrophages but not destroyed. The macrophages with their now protected residents become encased in clusters or granulomas. Since the bacteria are not actually destroyed, if the granulomas are disrupted, an activated infection can resurface. This explains why the tuberculosis bacilli, which are resistant to digestion by macrophages and therefore protected from antibiotic treatment, can be so difficult to treat. It is believed that some mastitiscausing organisms behave in an analogous manner. Specifically, such organisms appear to find "protected" areas (likely walled off by connective tissue elements) so that they are safe from destruction. This could explain why some cases of clinical mastitis are repeatedly caused by the same organism as well as reoccurring cases of Subclinical mastitis.Some of the important substances involved in inflammation and their effects are summarized in Table 15.1. Before we move to specific immunity, let's consider some of the effects of these molecules and related molecules involved in inflammation and chemical defenses. Some tissue proteins can inhibit or slow bacterial cell growth. For example, lactoferrin is a protein that is produced in secretions of the nonlactating mammary gland, where it accumulates to a high concentration. This is important because is acts to bind iron that is needed for bacterial cell growth (Brock, 2012). Histamine, released by local mast cells at the site of an attack, increases capillary permeability as well as vasodilation so that blood flow is increased. Mast cells also produce molecules that induce chemotaxis, which act as signals to induce the migration of phagocytic cells. Moreover, the binding of these agents to receptors on the cell surface of macrophages increases Ca++ flow into the cells. The increase in Ca++ activates contractile elements in the cells that allow ameba-like movement of the cells along the gradient of chemotactic signals (Box 15.1).
As illustrated in Figure 15.3, kallikrein secreted from phagocytes cleaves inactive Idninogens produced by the liver. These molecules appear in circulation and
Box 15.1 Protective secretions
As mentioned in an earlier chapter, it is believed that secretions from skin glands evolved to provide moisture and protection (antimicrobial agents) for the parchment-like shelled eggs laid by mammalian ancestors. Evolutionary pressure likely promoted the incorporation of molecules such as lysozyme or iron-binding lactoferrin into these secretions. Certainly, the capacity to prevent desiccation of the eggs and protection from microbial attack would have been highly beneficial. The importance of lactoferrin in particular has come full circle with the production of human-specific lactoferrin in milk of transgenic animals as well as the appearance of milk-extracted lactoferrin in products such as infant formula, human and pet foods, skin care products, toothpaste, and mouthwash (Stelwagen et al., 2008; Brock, 2012).
| Table 15.1. Agents involved in inflammation. | ||
| Agent | Source | Reaction |
| Histamine | Basophils and mast cells; released after injury, response to microorganisms or secretions from neutrophils | Vasodilation Ofarterioles increased permeability |
| Prostaglandins | From arachidonic acid present in cell membranes of neutrophils, platelets, and so on | Stimulation of endothelial cells and leukocytes to secrete inflammatory modulators |
| Kinins | Kininogens into interstitial fluid cleaved by kallikrein to produce active peptide | Effects similar to histamine but also chemotaxis |
| Complement | System of ~20 plasma proteins | When activated mechanism to inactivate bacteria and target foreign debris for removal |
| Cyokines | Circulation, lymphocytes, macrophages, and other immune cells | Examples include interferons, interleukins, and others all act to regulate immune function |
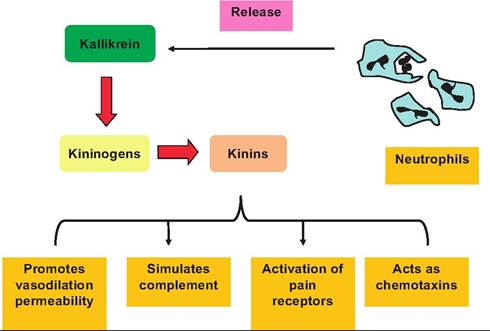
Fig. 15.3. Role of kallikrein in inflammation. Activated neutrophils release kallikrein which acts enzymatically on kininogens to produce kinins. These kinins promote multiple reactions that ultimately aid the body to fight inflammation.
diffuse into inflamed areas because of increased vasodilation, where they are converted into active kinins. Kinins subsequently stimulate several steps in complement activation, reinforce vascular bed changes initiated by histamine release, activate pain receptors, and serve as chemotactic agents to produce even more leukocyte migration and more kinin production in a positive-feedback pattern of action. This enhances the opportunity to defeat the offending agent. These inflammatory reactions can be likened to an alarm response as a large mixture of chemicals is released into the extracellular fluids.
Macrophages are especially important because these cells (along with certain epithelial cells in mucosal membranes) express proteins on their surface called Toll-Iike receptors (TLR). The receptors were named because they have a similar structure to a protein transcribed by the Toll gene in drosophila. They are also characterized as pattern recognition receptors and are appreciated for their capacity to recognize or bind molecules that are present on many pathogens but distinct from host surface molecules. These groupings of molecules are called pathogen-associated molecular patterns or PAMPs. The TLRs combined with Interleukin-I receptors are part of super family of receptors with similar attributes based on the fact that all of its members share the Toll-ILreceptor structural domain.
Cytokine is a general term given to the chemical messengers that regulate cells involved in immune responses. However, based on genetic linkages and structural similarities, many hormones and growth factors can be classified as cytokines. Interleukins are a subclass of cytokines that were first identified because they are secreted by leukocytes. At least 36 distinct interleukins are known. Most are produced by helper T lymphocytes, which are identified from their actions and expression of specific cell surface markers, in particular CD4. The interleukins are especially significant in the induction and differentiation of T cells, B cells, and other hematopoietic cells. It is telling that interleukins are also detected in other nonimmune tissues, and receptors for these proteins are widely distributed. Thus, roles for these powerful signaling molecules will certainly expand in the future.
At the present time, 13 distinct TLRs have been identified. Each of the receptors recognizes a particular class of microbes based on the components of the bacterial cell wall. Activation of the TLR in turn triggers the release of cytokines. Most cytokines are low molecular weight, soluble proteins produced following activation of a "sensing" cell (e.g., the macrophages with activated TLR as described earlier) (Rawlings et al., 2012).
Cytokines are produced by nearly all cells involved in immunity, but the T cells are especially important. The activation of cytokine-producing cells triggers them to synthesize and secrete their particular class of cytokines. The cytokines then bind to specific cytokine receptors on other cells of the immune system and influence their activity in some manner. Actions of cytokines are described as multifaceted, redundant, and pleiotropic. The idea is that a specific cytokine is likely to impact multiple target cells, not just one type of cell. In addition, some cytokines are antagonistic so that one cytokine stimulates a particular action while another cytokine inhibits the same function. In other cases, cytokines act synergistically to produce a greater effect than either would have alone. These features mean that there are enormous opportunities for control and regulation of immune responses. Most of the cytokines act on leukocytes or the endothelial cells to affect inflammatory responses. But there is also overlap between classic hormones and cytokine signaling pathways. For example, leukocytes and macrophages express prolactin receptors. It seems likely that, in addition to its well-characterized actions in mammary development and reproduction, prolactin also acts as a cytokine in the immune system. Leptin, the hormone produced by adipocytes, is also now recognized for its impacts on the immune system (Procaccini et al., 2012). Some features of a few selected cytokines are described below.
Tumor necrosis factor a (TNFoc) is the major cytokine involved in acute inflammation. It is primarily synthesized by monocytes, macrophages, and helper T cells. When produced in very large amounts, it is believed to be the cause of systemic shock in severe reactions. It acts on the endothelial cells to stimulate inflammation and blood clotting. It also promotes endothelial cells to secrete selectins (adhesion molecules) that are important for diapedesis of leukocytes. Furthermore, it triggers macrophages and endothelial cells to secrete chemokines. Chemokines also impact diapedesis and chemotaxis, and additionally promote macrophages to secrete interleukins (see later discussion). Finally, TNFoc is directly cytotoxic for some tumor cells, thus explaining its name.
The interleukins include a very large family of structurally similar cytokines. Interleukin 1 (IL-I) is especially significant. It has actions functionally similar to those of TNFoc (an example of some of the redundancies among cytokines). Common effects include induction of fever and sleep and stimulation of collagen synthesis as well as collagenase needed for tissue repair and remodeling. It also stimulates T and B lymphocytes to proliferate. IL-I is also produced by monocytes, macrophages, dendritic cells, and a variety of other cells in the body.
Other interleukins include IL-2 that is secreted primarily by helper T cells to stimulate the proliferation of helper T cells and activate the natural killer T cells. It is also called T cell growth factor. IL-3 promotes hemopoiesis to generate precursors of lymphocytes and mast cells. IL-4, also produced by the helper T cells, stimulates B cells and enhances antibody secretion by active plasma cells, especially secretion of IgE. IL-5 behaves in a similar fashion but is more likely to promote plasma cells to secrete IgA-type antibodies. It also acts as a chemoattractant for eosinophils. IL-6 has wide-ranging effects, including promotion of differentiation of B cells into plasma cells and stimulation of the liver to secrete a mannose-binding protein that triggers complement protein binding to the surface of microorganisms that have mannose-containing polysaccharides in their cell walls. IL-8 promotes angiogenesis, an action that is clearly important in repair of tissue damage. IL-IO acts to dampen or turn down immune responses, so it is important in overall regulation of immune function.
Transforming growth factor β (TGF-β) is also a suppressor of immune responses by its capacity to inhibit the proliferation and function of T cells and proliferation of B cells. Along with the other cytokines released, TGF-β is an important participant because of its role in several stages of wound healing. Vascular endothelial cells are early responders. There is enhanced secretion of adhesion molecules (VCAM-1, ELAM-1, ICAM-1) in the area of the endothelial cells that gives a foundation for the anchoring of circulating leukocytes that express receptors (integ- rins, selectins, etc.) to recognize these adhesion factors and allow accumulation of the leukocytes, chemotactic attraction, and diapedesis. Indeed, TGF-β is a potent chemoattractant in its own right. Because of its effects on secretion of extracellular matrix proteins by stromal cells (fibroblasts), it also promotes tissue repair. TGF-β is produced by T lymphocytes, macrophages, and other stromal tissue cells and appears in circulation in a latent form that is activated by tissue proteases.
The colony-stimulating factors (CSF) are an additional group of proteins that impact immune function by their ability to induce production of colonies of the different leukocyte types in the bone marrow. Some specific CSF members include granulocyte macrophage colony-stimulating factor (GM-CSF), granulocyte colony-stimulating factor (G-CSF), and macrophage colony-stimulating factor (M-CSF). Aside from effects on proliferation, the CSFs also influence leukocyte function. For example, when GM-CSF binds to receptors on neutrophils, eosinophils, or monocytes, it activates the cells and also enhances survival of the cells. GM-CSF increases the capacity of these phagocytes to form pavements involved in diapedesis between endothelial cells and improves the ability of the cells to destroy engulfed bacterial cells. CSFs are produced mostly by T cells and macrophages.
A final group of cytokines to introduce are the interferons. As the name suggests, these molecules are characterized by having an ability to interfere with something. In this case, it is the replication of viruses. Like other cytokines, there are multiple types of interferons (α, β, and γ). In short, as illustrated in Figure 15.4, interferons provide some resistance to viral infections by interfering with the replication of the virus in neighboring potential host cells. In addition, interferons enhance the phagocytic activity of macrophages and stimulate the secretion of antibodies by plasma cells. Furthermore, these molecules markedly improve the function of natural killer and cytotoxic T cells, which are important in the destruction of virus- infected and cancerous cells.
Figure 15.5 provides an overview of an essential aspect of inflammation, the migration of phagocytes to the affected area. In this example, the rapid response of polymorphonuclear (PMN) leukocytes into the mammary gland following an experimental insult (i.e., intramammary infusion of endotoxin) is shown. Since the site of the inflammation is deep within the areas of the mammary gland that store milk, the phagocytes must move out of the capillaries (between the endothelial cells) but additionally pass the basement membrane and between the epithelial cells that compose the outer structure of the mammary alveoli. In these cases, the phagocytes have responded to chemoattractant agents in milk that have diffused into surrounding interstitial fluids and local capillary beds.
As an example, the somatic cell count of raw milk is the most common dairy producer-related method to
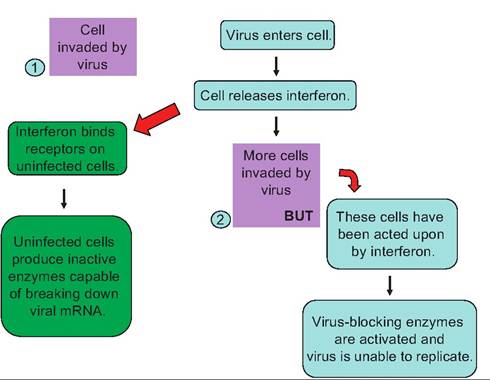
Fig. 15.4. Action of interferon. In step 1, a cell is invaded by a virus. In response the infected cell produces and secretes interferon. The interferon binds to receptors on neighboring cells and induces the cells to produce inactive enzymes capable of breaking down viral rr>RNA. In step 2, a cell previously impacted by interferon is subsequently invaded by the virus. The virus blocking enzymes are activated, and viral replication in this cell is prevented.
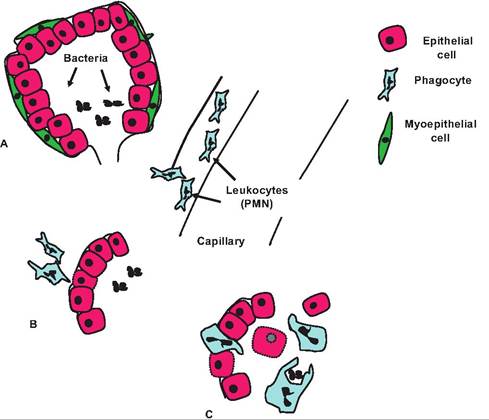
Fig. 15.5. Leukocyte (PMN) response in the mammary gland. Chemotaxis causes PMNs to migrate from the capillaries and form a pavement along the basement membrane of the infected alveoli (A). Some groups of epithelial cells become damaged by the toxins or PMN activity and are sloughed into the alveolar spaces (damaged cells indicated by the broken cell membranes). Neutrophils and macrophages accumulate (B) where they engulf bacterial cells (C) and destroy them along with cell debris (Nickerson and Heald, 1981; Capuco et al., 1 986).
evaluate milk quality and udder health status of individual lactating cows. Leukocytes and a small percentage of epithelial cells normally occur in milk. This combination of cells is referred to the milk somatic cell count (MSCC). The term somatic, which means body, alludes to the fact that these are normal body-derived cells. Most (~98%) of the cells are leukocytes, and most of these are neutrophils, sometimes called PMN leukocytes. This descriptive term is a reference to the Iobed nucleus of these cells. Milk from uninfected cows typically contains fewer than 200,000 cells per mL, and it is not uncommon to find uninfected cows with MSCC of 50,000 cells or fewer. Milk samples with values more than 400,000 cells per mL are very likely from cows with inflammation most likely caused by mastitis-producing organisms. These leukocytes enter the milk as a consequence of homing to the mammary gland from the bloodstream in response to chemicals released directly by bacterial cells or materials released by injured mammary cells. These chemicals induce chemotaxis that initially recruits neutrophils and thereafter macrophages (monocytes) into the udder. Since an increase in MSCC is closely correlated with intramammary infection, the MSCC is measured for milk samples collected as a part of routine monitoring of milk composition in many dairy herds. However, it is important to remember that, strictly speaking, bacteria-induced mastitis can only be confirmed by the isolation of pathogenic organisms in aseptically
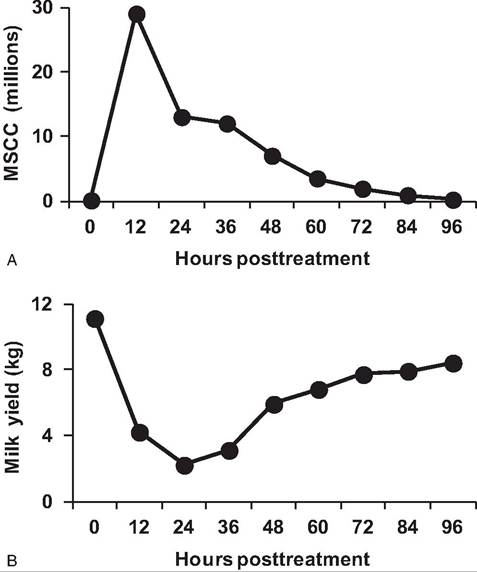
Fig. 15.6. Cell and milk production responses in cows. Changes in MSCC (upper panel) and milk production (lower panel) in cows given an intramammary infusion of bacterial endotoxin are shown. Note the marked increased and corresponding decrease in mammary function. Adapted from McFadden et al. (1988).
collected milk samples by approved bacteriological methods. Figure 15.6 shows the marked ability of PMN to respond to an intramammary signal. In this case, the mammary gland of the cow was infused with purified endotoxin diluted in sterile physiological saline. By the time of the next milking, the MSCC had increased ~30-fold, and there was a corresponding sharp decrease in milk production. However, after several days, the cell response and milk production returned to normal. This is an experimental situation, but it certainly demonstrates the dramatic response that phagocytic cells can make in response to stimulation.
Fortunately, most tissue regions have macrophages that are residents. For example, with a skin break, these resident macrophages can begin phagocytizing microorganisms (assuming they are recognized as invaders) almost immediately. However, the number of cells is limited so that a full-blown response depends on chemotaxis to recruit additional phagocytes (more macrophages and neutrophils). It is important, of course, that these cells express the ability to recognize foreign material so that normal cells are not harmed. The tagging of materials to be engulfed is complex, but when it comes to bacterial cells, the complement system is especially important. This is because the effects of activated complement can be very potent, but secondly, the presence of specific antibodies on the surface of the bacterial cells (or other materials) acts to mark these invaders for destruction by phagocytes as well as attack by complement proteins.
The complement system, usually simply called complement, is a complex of at least 20 proteins present in the circulation (originally produced by the liver). These proteins provide an important mechanism for the destruction of foreign substances because, when activated, they greatly enhance the inflammatory response, but, even more impressively, they can stimulate the direct destruction of bacteria and some other cells by causing the cells to rupture. Aside from direct effects on bacterial cells, activated complement also enhances inflammation in several ways:
• stimulates release of histamine from mast cells
• promotes vasodilation and thereby increased vascular permeability
• activates kinins
• coats the surface of microorganisms and act as opsonizing agents.
Thus, while complement is really nonspecific, it clearly enhances or "complements" the immune response, hence the name. The effects of complement on bacterial cells are illustrated in Figure 15.7.
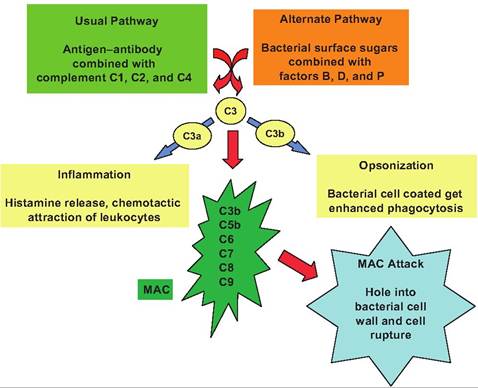
Fig. 15.7. Complement activation. In the classic activation pathway the binding of antibody to bacterial antigens induces complex formation with complement proteins C1, C2, and C4. Alternatively, plasma proteins (factors B, D, and P) can bind with surface polysaccharides in the cell wall of some bacteria and fungi. Regardless of the pathway, both converge with the activation of complement protein C3. This activation produces a cleavage reaction to yield C3a and C3b. When C3b binds to the bacterial cell surface, it initiates the recruitment Ofcomplement proteins C5b, C6, C7, C8, and C9 to produce the membrane attack complex (MAC). This complex penetrates the cell wall to create a hole or pore that leads to cell lysis. In addition, binding of C3b induces opsonization or coating of the bacterial cell which enhances targeting by phagocytes for engulfment. Finally, the presence of free C3a and C5a in the area promotes inflammation.