Structure and function of DNA and RNA
As we have now learned, essentially all of the biochemical reactions to produce energy or build cellular
id="Picutre 87" class="lazyload" data-src="/files/uch_group75/uch_pgroup311/uch_uch7424/image/image087.jpg">
Fig.
3.21. Amino acid nitrogen metabolism. A general overview of nitrogen flow in amino acid metabolism is shown. Transamination reactions provide a mechanism to Catabolize a variety of amino acids by transfer of the amine group to pyruvate (alanine transaminase, i.e., alanine is produced from pyruvate in the transfer) or transfer of the amino acid amine group to a-ketoglutarate catalyzed by the enzyme glutamate transferase; that is, glutamate is produced by the transfer. The ability to generate glutamate is especially important because it is the only amino acid in most mammals that readily undergoes oxidative deamination. This provides a mechanism to excrete the amine group in the form of ammonia, which is typically converted to the less toxic urea.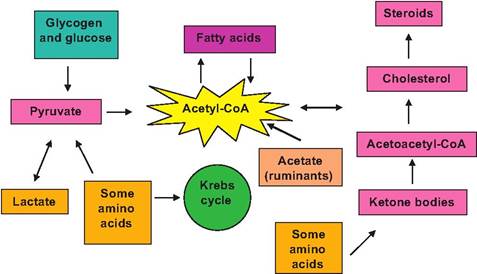
Fig. 3.22. Overview of acetyl-CoA metabolism.
components depend on enzymes. Other proteins are critical components of various cellular organelles. This means it is nearly impossible to overestimate the importance of proteins. It follows then that the creation and activation of proteins must be carefully orchestrated so that the suitable enzymes are available when required and the proteins for organelle generation are present. In many respects, cellular functioning follows the now-popular business concept of on-time
delivery. This ultimately goes back to regulation of gene expression and controls that affect transcription and translation (Chapter 2). Although we have considered some of the basics of RNA and DNA structure, we will now review protein synthesis in a bit more detail. It is reasonable to think of cells as miniature protein factories and that the particular combination and number of proteins fashioned determine the functional attributes of the cell.
For example, although all cells have common components, it is logical to predict that the complement of proteins needed for adequate functioning of a fibroblast would be very different from the complement of proteins made by a secretory epithelial cell from the pancreas.Protein synthesis
DNA not only provides the template to direct its own replication, it also provides the blueprint for the synthesis of proteins by its ability to direct the formation of mRNA. As you should recall, this process is called transcription. Figure 3.23 gives an overview of information transfer from DNA to RNA to proteins for a eukaryotic cell. An important aspect for eukaryotic cells compared with prokaryotic cells is the fact that the genes of higher organisms are interrupted by DNA sequences that do not code for the ultimate protein product. DNA sequences that do correspond with the protein are called exons and those that make the intervening sequences are called introns. A single gene may contain 50 or more introns. Moreover, these introns range from as few as 60 to more than 100,000 nucleotides. This means that the initial RNA strand must be processed to remove the introns before it can be used to accurately direct protein synthesis. One of the great puzzles of cell biology is to understand the physiological significance of the introns. On the surface it seems wasteful for the cell to spend the biochemical resources to synthesize introns initially and then to recreate segments of the mRNA molecule that must be removed and discarded.
Proteins are chains of amino acids. Genes can be viewed as the segment of DNA that provides the directions for construction. There are also a few genes that direct the creation of specialized variants of RNA, but certainly, most genes direct protein synthesis. The nucleotide bases A, T, C, and G make up the foundation of the code for these assembly instructions. A sequence of three bases or triplet specifies a particular amino acid. For example, the sequence AAA indicates the amino acid lysine.
This means that the sequence of triplets in the mature processed mRNA that leaves the nucleus (see Fig. 3.23) spells out the exact sequence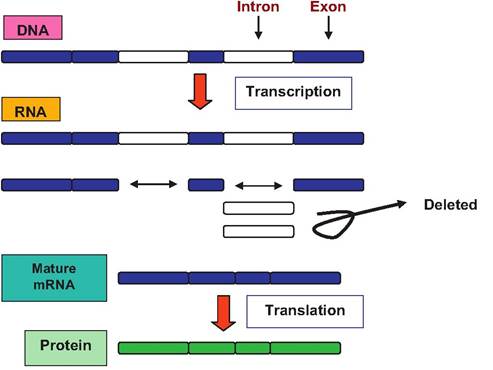
Fig. 3.23. Overview of transcription and translation and protein synthesis.
of amino acids for a particular protein. These triplet sequences are the genetic code.
So what are the mechanics of protein synthesis? This requires the combined activities of three types of RNA molecules: mRNA, rRNA (ribosomal), and tRNA (transfer RNA). The ribosomes are the cellular organelles that are the focus of actual protein synthesis. These organelles are composed of subunits derived from RNA and protein molecules and are fabricated by the nucleolus inside the nucleus of the cell. To use an automobile analogy, rRNA can be thought of as the factory that houses and organizes the needed components. Processed mRNA provides the blueprint for protein assembly. It could be thought of as the assembly line that is organized along the factory floor. The tRNA can be thought of as the forklifts and cranes that bring the components (in our case amino acids) needed to make the car (protein in our case). Of course with cars, the components are welded or bolted together by workers or workers driving machinery. In our analogy, as we have seen with other biochemical processes, the bolting or welding of the amino acids (peptide bond formation) requires energy supplied by ATP hydrolysis and is accomplished by enzyme activity. To summarize, polypeptide synthesis requires two fundamental steps: (1) transcription, during which time the DNA "information" is encoded into mRNA, and (2) translation, the process by which the "information" in mRNA is decoded and is used to manufacture the proteins. This process is outlined in Figure 3.24.
Let us begin with detailing transcription. The first step involves the unwinding or unraveling of a segment
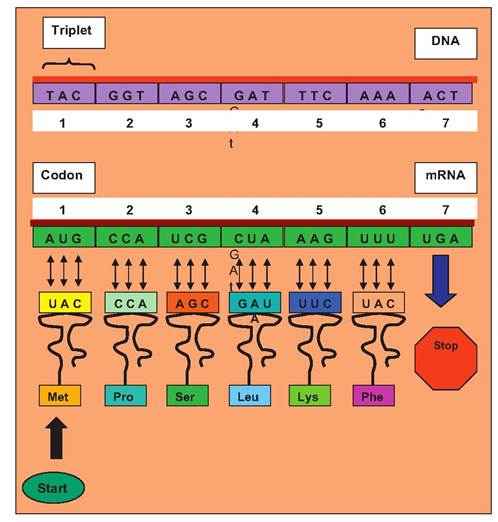
Fig.
3.24. Information transfer from DNA to RNA. Processed mRNA is utilized to direct the fabrication of the growing protein chain via transfer RNA and their attached amino acids which have complementary binding (anticodon) to the codons of the mRNA.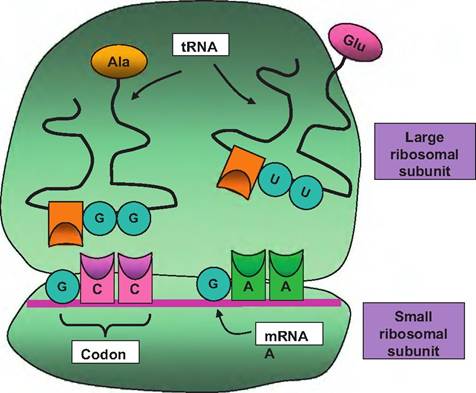
Fig. 3.25. Transfer RNA. Transfer RNA transports amino acids (alanine and glutamic acid) in this example, to the ribosome for interaction with the mRNA. The triplet codon of the mRNA stand corresponds with complementary binding of the anticodon of the tRNA. Following binding of adjacent amino acids, the enzyme aminoacyl-tRNA synthetase catalyzes the formation of peptide bonds. As the mRNA passes through the ribosome complex, the protein chain continues to elongate until a stop codon is reached and the nascent protein chain is released.
of DNAthat is destined for use. The cytoplasm contains specialized molecules (some of which are mediators of hormone action, by the way) called transcription factors. These molecules have the capacity to bind to regions near the beginning or "start" sequence of a gene. This region, which is not part of the final mRNA product, is called the gene promoter. In simplistic terms, activation of this region sets in motion or promotes the subsequent transcription of a particular gene. A great deal of cellular activity is determined by the complex of particular transcription factors that are unleashed in a cell at a given moment and which promoter regions are available to be acted upon.
The transcription factor acts to mediate the binding and initiation of the enzyme RNA polymerase. RNA polymerase allows opening of the double-stranded DNA. One strand then serves as the template to produce the complementary mRNA molecule (sense strand). For example, if the triplet sequence were AGC, the corresponding mRNA would be UCG. You might recall that RNA differs from DNA in that it is single-stranded, contains ribose instead of deoxyribose, and the base uracil (U) substitutes for thymine (T).
This explains the U instead of T in the newly created RNA triplet. The strand of DNA that is not used as a template is called the antisense strand. Each triplet of the DNA corresponds with a three-base sequence of the mRNA called a codon. Since there are four different nucleotides in RNA or DNA, there are 43 or 64 possible codons. Three of these (UAA, UAG, and UGA) serve as stop signals and AUG, which codes for the amino acid methionine, serves as a start signal. Since there are only 20 common amino acids, some amino acids are coded by more than one triplet sequence.Once the mature or processed mRNA (introns removed) reaches the cytoplasm, it joins the smaller of the ribosomal subunits. At this point the tRNA comes into action. The tRNA binds its amino acid and transports it to the ribosome. There are approximately 20 different tRNA variants, which correspond with each of the different amino acids. Not only must the tRNA recognize its particular amino acid (by possessing a unique binding site), it must also recognize the triplet codon of the mRNA molecule. These tRNA molecules have a complex wire hanger-like conformation that allows simultaneous recognition of the mRNA codon by complementary base pairing (anticodon) along with orientation of the attached amino acid into position to allow the enzyme aminoacyl-tRNA synthetase to catalyze formation of peptide bonds between adjacent amino acids attached to the mRNA (see Fig. 3.25).
This is accomplished because of the elegant structure that is produced by the combination of the large and small ribosomal subunits attached to the mRNA molecule. There is a binding site for the mRNA and three sites for binding of the tRNA molecules. The charged tRNA (one that has its amino acid attached) binds to the A (attachment) site. As the peptide bonds are formed, there is a shift to the P site, which holds the growing peptide chain. As newly charged tRNA molecules arrive at the ribosome, the old empty tRNA shifts to the E (exit) site and is released to capture another amino acid.
As the mRNA codons are progressively read, the mRNA passes through the ribosome as the elongating amino acid chain appears. As the mRNA emerges, other ribosomes can attach and begin the process of translation. This means that many protein molecules can be fabricated simultaneously. These complexes are called polyribosomes. This is illustrated in Figure 3.26. The mRNA strand continues to be read, and the protein chain grows until its stop codon is reached. At this point, the elongating protein chain and the ribosomal subunits is freed from the ribosome.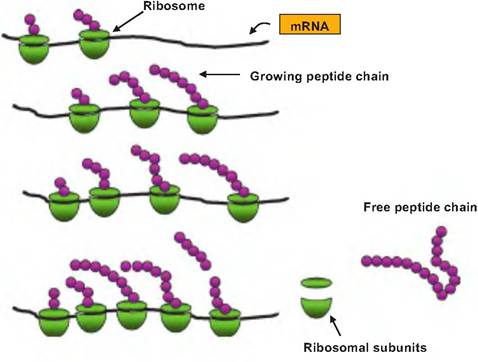
Fig. 3.26. Development of polyribosomes. Each polyribosome consists of a strand of mRNA that is being transcribed by multiple ribosomes. As the mRNA moves through the ribosome, each codon is sequentially read, and peptide bonds are formed between adjacent amino acids. Ribosomes that have been attached longer have the correspondingly longer peptide chains. This progression (left to right) continues until the stop codon is reached when the nascent peptide is released along with the ribosomal subunits.
Chapter summary
Just as we have all heard the expression, "it is all about the money," physiologically speaking, it is all about thermodynamics and managing to convert substrates into macromolecules and macromolecules into cell structures. Cellular biochemistry concerns the myriad of conversions and interactions necessary to acquire building blocks and the energy to perform critical anabolic and catabolic reactions. Only a portion of the energy available from the breaking of the chemical bonds in molecules from ingested feed materials can be captured. Moreover, the balance of energy captured, lost, and needed for maintenance ultimately determines the net energy remaining to provide for lactation, growth, work, egg production, or hair growth in our animal systems.
Cellular energy revolves around ATP and the ability of cells to utilize energy that becomes available with the breaking of the bonds that attach the third phosphate group in the molecule. ATP is composed of adenosine + ribose + three phosphate groups coupled in sequence to carbon 5 of the ribose backbone. Removal of phosphate groups in succession would lead from ATP to ADP to adenosine monophosphate (AMP). Most cells utilize the catabolism of the hexose sugar glucose to generate needed ATP
The initial processing of glucose by the cell is called glycolysis (translated as the breaking or lysis of glucose). The steps in this biochemical pathway take place in the cytoplasm of the cell because the necessary enzymes are produced by the cell and are maintained within the cytoplasm. Movement of glucose into the cells requires a transporter or carrier. Importantly, once in the cytoplasm, glucose is phosphorylated, that is, converted into glucose-6-phosphate by the enzyme hexose kinase. This step is important because the addition of the phosphate group prevents the glucose from interacting with the membrane carrier. Thus, the facilitated diffusion of additional glucose into the cell is not impaired. A series of enzyme-mediated reactions ultimately leads to the generation of two molecules of the 3-carbon intermediate pyruvate for each glucose molecule that enters the cycle. In the absence of oxygen, pyruvate is converted to lactate. This is important because the conversion to lactate via the enzyme lactate dehydrogenase is coupled with the conversion of NADH to NAD. NAD is necessary for continued operation of glycolysis. There is a net gain of two ATP under anaerobic conditions.
With the availability of oxygen, pyruvate from glycolysis enters the mitochondria to participate in the Krebs cycle as it is decarboxylated to create acetyl- CoA. With each turn of the Krebs cycle, multiple molecules of reduced NAD and FAD are produced. These coenzymes enter the electron chain in the mitochondria where a series of coupled electron transfers provide the power to generate ATR The final acceptor of electrons is oxygen. This ultimately explains why oxygen is essential. In the absence of oxygen, the electron chain transfers grind to a halt. In total, for each molecule of glucose through both glycolysis and Krebs cycle, 38 ATP are generated.
The importance of these two pathways cannot be overstated. Consequently, it is critical that the supply of glucose (or molecules that can participate in the Krebs cycle) is maintained. Glycogenesis refers to the conversion of excess glucose into glycogen (animal starch). Muscle and liver cells are primary sites for storage of glycogen. Gluconeogenesis refers to the conversion of other nutrients into glucose. These processes are particularly critical in ruminants because nearly all available dietary glucose is fermented into volatile fatty acids (acetate, butyrate, and propionate) by rumen bacteria. Fortunately, the liver of ruminants is able to convert propionate into glucose to supply tissues and functions (brain and lactose production) that have absolute requirements for glucose. For cows and other ruminants, acetate from rumen fermentation can be utilized as an energy source (ATP generation) after it is converted into acetyl-CoA in the mitochondria.
Glucose and carbohydrate metabolism is essential, but there are limited stores of glycogen. Instead, long-term energy storage relies on sequestration of triglycerides in adipocytes. Lipogenesis is the making, packaging, and storage of these triglycerides. When there is a call for release of FFAs into the blood, this is called lipolysis. FFAs are oxidized via a biochemical pathway called β-oxidation. This process efficiently produces multiple molecules of acetyl-CoA for use in the Krebs cycle. For example, a molecule of the 16-carbon fatty acid oleic produces a net of 135 molecules of ATP
Several different amino acids can also be deaminated to produce intermediates of the Krebs cycle and thus also be used for ATP production. The subsequent combination of amine group from the catab- olized amino acid with CO2 produces urea which is much less toxic than ammonia.
Review questions and answers are available online.