The heart
Anatomy of the heart
Location and exterior landmarks
The heart is an inverted cone-shaped structure located in the mediastinum, a mass of tissue occupying the medial region of the thoracic cavity extending from the sternum to the vertebral column, and between the lungs.
The apex, or "pointed" end of the heart, is directed Caudoventrally; the base, or top of the heart, is directed dorsocranially (Fig. 13.6).The cranial and caudal sides of the heart can be located by other structures. The auricles point left, with the pulmonary trunk located between the two auricles. The aortic arch projects caudally.
The coronary groove partially encircles the heart except at the conus and indicates the separation of the atria and ventricles. The conus is the pyramidal structure that is the right ventricular outflow tract into the pulmonary trunk. The interventricular grooves indicate the divisions between the two ventricles. The two auricles are visible on the left side of the heart, with the pulmonary trunk between them.
Pericardium
The membrane surrounding the heart is the pericardium. It consists of the fibrous pericardium and serous pericardium. The fibrous pericardium is a tough, inelastic, dense, irregular connective tissue sac with one end attaching to the diaphragm and the other open end fusing with the connective tissue surrounding the blood vessels entering and leaving the heart. The fibrous pericardium anchors the heart within the mediastinum and prevents overfilling of the heart. Inside the fibrous pericardium is the serous pericardium, consisting of a parietal and visceral layer. The parietal layer lines the internal surface of the fibrous pericardium; the visceral layer, also called the epicardium, is an integral part of the heart wall.
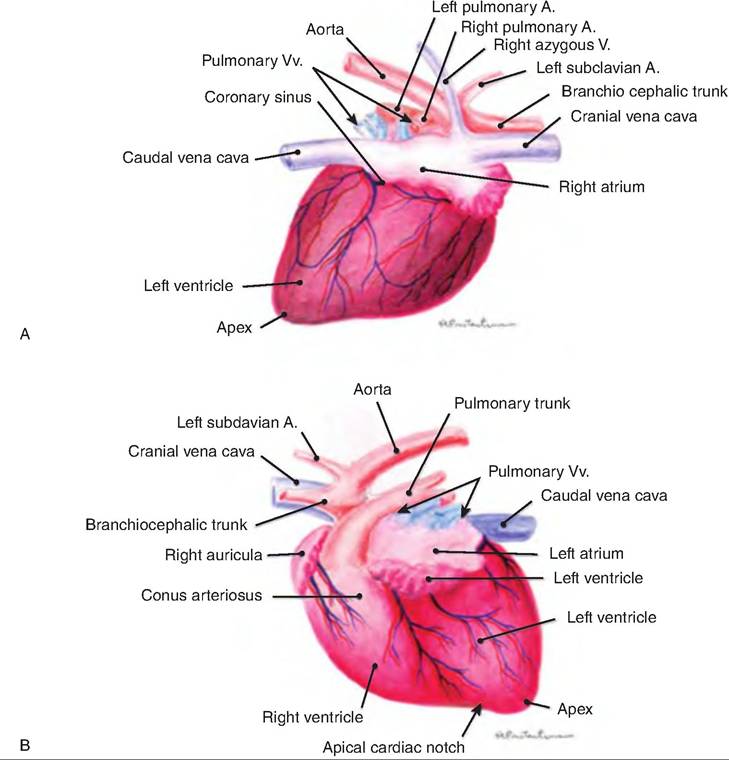
Fig.
13.6. External structure of the heart. (A) Atrial side of the cat heart. (B) Auricular side of the cat heart. A, artery; v, vein. (Reprinted from Constantinescu, 2002. With permission from the publisher.)Inflammation of the pericardium is called pericarditis. This results in decreased production of serous fluid, and a roughened serous membrane. As a result, the beating heart can be heard with a stethoscope rubbing against the serous layer (pericardial friction rubs). In severe cases, inflammation leads to excess fluid production, which compresses the heart and decreases its pumping ability.
Layers of the heart
The heart wall consists of three layers: epicardium, myocardium, and endocardium. The epicardium is the outermost layer, and it is the visceral layer of the pericardium. It consists of a thin, transparent layer of mesothelium and connective tissue. The middle layer, or myocardium, is cardiac muscle and makes up the bulk of the heart. The innermost endocardium is a thin layer of connective tissue providing a smooth lining for the chambers of the heart and valves. The endocardium is continuous with the endothelial lining of the large blood vessels attached to the heart.
Cardiac muscle is also called involuntary, striated muscle. Like skeletal muscle, it contains actin and myosin that is organized into sarcomeres.
Fibrous skeleton of the heart
The heart also contains dense connective tissue surrounding the valves, forming a fibrous skeleton. In addition to forming a point of attachment for the valves, the fibrous skeleton serves to eclectically insulate the atria from the ventricles (Box 13.5).
Heart chambers and vessels
The heart has four chambers. Two atria located superiorly receive blood and pump it to the ventricles. Two ventricles located posteriorly pump the blood away from the heart (Fig. 13.6). The atria are separated by the interatrial septum; the ventricles are separated by the interventricular septum. There is an oval depression on the interatrial septum called the fossa ovalis (Fig.
13.7), a remnant of the foramen ovale, which is an opening between the atria in the fetus that closes shortly before birth.Box 13.5 Dilated cardiomyopathy
Dilated cardiomyopathy (DCM) is a disease characterized by dilation or enlargement of the heart chambers resulting in an abnormally large heart. This disease eventually results in heart failure, since the damaged heart muscle is too weak to efficiently pump blood to the rest of the body. DCM is very common in dogs, representing the most common reason for congestive heart failure (CHF). The left ventricle is most always involved. Since the myocardium cannot work effectively to pump blood out of the heart, subsequent backup of blood into the left atrium and ultimately into the lungs occurs commonly. This backup of blood into the lungs results in pulmonary edema and is a sign of congestive heart failure.
The treatment of dogs with dilated cardiomyopathy varies with the severity of heart failure and specific organ damage. Treatment may include oxygen administration, fluid therapy, and administration of drugs that improve breathing (bronchodilators) and drugs that modify heart function, such as control of the arrhythmias. If low doses of anti-arrhythmic drugs are effective, the heart can often be stabilized. Serious ventricular arrhythmias that can only be controlled by high doses of antiar- rhythmic drugs have a poorer prognosis.
Atria
The atria are the receiving chambers of the heart. Protruding from the atria are the auricles, which increase the atrial volume. The auricles are lined with pectinate muscles, making them appear as if they were raked with a comb. The atria are relatively small and thinwalled, since they need to pump blood only to the ventricles.
Blood enters the right atrium from three veins: (1) the superior vena cava returns blood from the body regions in front of the diaphragm, (2) the inferior vena cava returns blood from areas posterior of the diaphragm, and (3) the coronary sinus collects blood draining the myocardium (Fig.
13.6). Blood passes from the right atrium into the right ventricle through the tricuspid valve, so named because it consists of three leaflets or cusps.Blood enters the left atrium via four pulmonary veins. Blood passes from the left atrium to the left ventricle via the bicuspid, or mitral, valve, named because it has two cusps.
Ventricles
The ventricles form the bulk of the heart. The right ventricle wall is thinner than the left since it has to
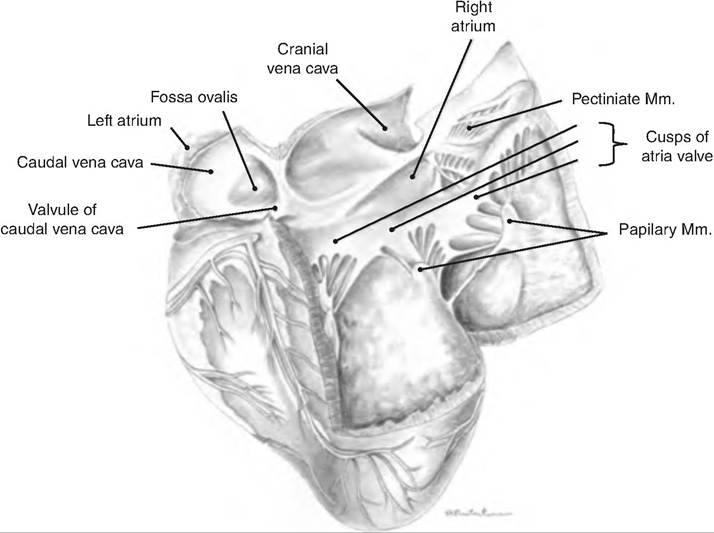
Fig. 13.7. Internal structure of the heart. The right side of the heart of a large ruminant is opened. Mm, muscle. (Reprinted from Constantinescu and Constantinescu, 2004. With permission from the publisher.)
Box 13.6 Feline dilated cardiomyopathy Hypertrophic cardiomyopathy (HCM) is a heart (cardio) muscle disease (myopathy) in which the muscular walls of the left ventricle thickens (hypertrophy). The left ventricular walls may hypertrophy secondarily to other diseases such as systemic hypertension, or the hypertrophy can be a primary disease in itself.
HCM is diagnosed when thickening of the left ventricular walls is not caused by another disease. As HCM progresses, it can alter the heart structure and impair its functioning in several ways, including the following: (1) Ventricular chamber size may be reduced, thus limiting its ability to fill with blood; (2) Ventricular wall stiffness usually increases, which impairs the ability of the ventricle to relax, preventing it from filling efficiently; (3) There may be an increase in ventricular pressure during relaxation (diastole), causing blood to back up into the vessels of the lungs and subsequent congestive heart failure, which includes pulmonary edema and/or pleural effusion (see vascular fluid into the lungs and/or pleural spaces).
Because the left ventricle is unable to fill adequately, less blood is pumped out to the body with each heartbeat. If the blood supply to other vital organs is inadequate, heart rate (HR) may increase as the body attempts to compensate.
A decrease in blood flow to the kidneys can result in an increased release of renin, which increases blood volume, increasing the pressures on the left side of the heart, and contributing to congestive heart failure.pump blood only through the lungs via the pulmonary trunk. The left ventricle pumps blood to the body via the aorta, the largest artery in the body.
Blood leaves the right ventricle via the pulmonary valve. The left ventricle forms the apex of the heart. Blood leaves the left ventricle via the aortic valve. During fetal development when there is no pulmonary respiration, there is a temporary blood vessel called the ductus arteriosus that shunts blood from the pulmonary trunk into the aorta. This vessel closes shortly after birth, leaving a remnant called the ligamentum arteriosum.
Inside the ventricles are muscle bundles called the papillary muscles, which serve as attachments for the chordae tendineae, tendinous cords attaching to the atrioventricular (AV) valves. The papillary muscles and chordae tendineae assist in valve function (Box 13.6).
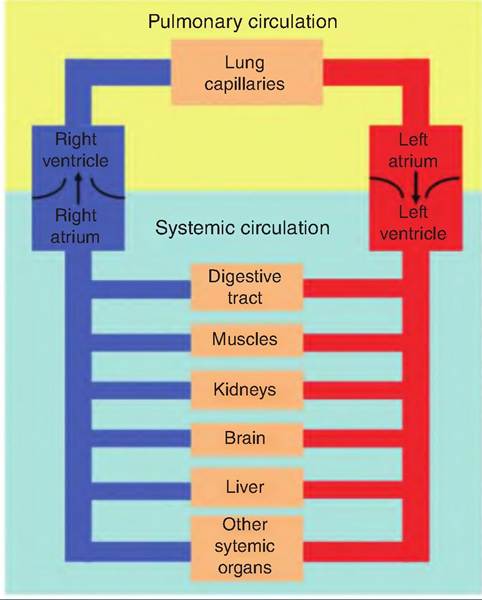
Fig. 13.8. Systemic and pulmonary circulation. The right side of the heart pumps deoxygenated blood to the pulmonary circuit, while the left side of the heart pumps oxygenated blood throughout the body.
Pathways of blood through the heart
The heart acts as two pumps, side by side. The pulmonary circuit carries blood to and from the lungs, and the systemic circuit transports blood throughout the remainder of the body (Fig. 13.8).
The right side of the heart receives deoxygenated blood from the body. This blood passes into the right atrium, through the tricuspid valve, and into the right ventricle. It is then pumped to the lungs via the pulmonary trunk. In contrast to other major arteries and veins in the body, the pulmonary artery carries oxy genpoor blood, while the pulmonary vein carries oxygenrich blood.
The left side of the heart receives freshly oxygenated blood arriving from the lungs via the pulmonary vein. The blood passes from the left atrium to the left ventricle via the bicuspid valve. Blood is then pumped from the left ventricle into the aorta, passing through the aortic valve.
Heart valve operation
Atrioventricular valves
The atrioventricular (AV) valves lie between the atrium and ventricles. When open, the cusps of the valves push into the ventricles, allowing blood to flow from the atrium to the ventricles. While the AV valves are open, the papillary muscles and chordae tendineae are relaxed. When the ventricles contract, pressure in the ventricles increases and pushes blood back toward the atria. This blood pushes the cusps of the valves back toward the atria, closing the valves. Simultaneously, the papillary muscles contract, pulling on the chordae tendineae. The chordae tendineae prevent the cusps of the AV valves from everting into the atria. Damage to the AV valves or chordae tendineae allows regurgitation of blood through the AV valves.
Semilunar valves
The semilunar valves include the aortic and pulmonary valves, which allow blood to pass from the ventricles into the aorta and pulmonary vein, respectively. These valves are made of three crescent-shaped cusps. As the pressure in the ventricles exceeds that in the arteries, blood passes from the heart into the arteries. As the ventricles relax, the backflow of blood catches the cusps and causes these valves to close, thus preventing the movement of blood back into the ventricles.
There are no valves located at the entrance of the venae cavae into the right atrium or pulmonary veins into the left atrium. So as the atria contract, a small amount of blood can backflow into these veins. However, contraction of the atria compresses the area where the veins attach, thus minimizing the backflow of blood.
Pulmonary, systemic, and coronary circulation
Pulmonary circulation
The pulmonary circulation transports deoxygenated blood from the right ventricle to the lungs where it picks up O2 while delivering CO2. The right side of the heart is responsible for the pulmonary circuit. Deoxygenated blood returning from the body enters the right atrium and passes into the right ventricle. The right ventricle pumps the blood into the pulmonary artery and into the pulmonary capillaries. Oxygenated blood is returned to the left atrium via the pulmonary vein.
Systemic circulation
The systemic circulation distributes oxygenated blood throughout the body. Blood is pumped from the left ventricle into the aorta, and then into smaller systemic arteries. These arteries give rise to arterioles that lead to systemic capillaries. Exchange of nutrients occurs across the capillary walls. Blood enters the systemic venules and then into the systemic veins that return the blood to the right atrium.
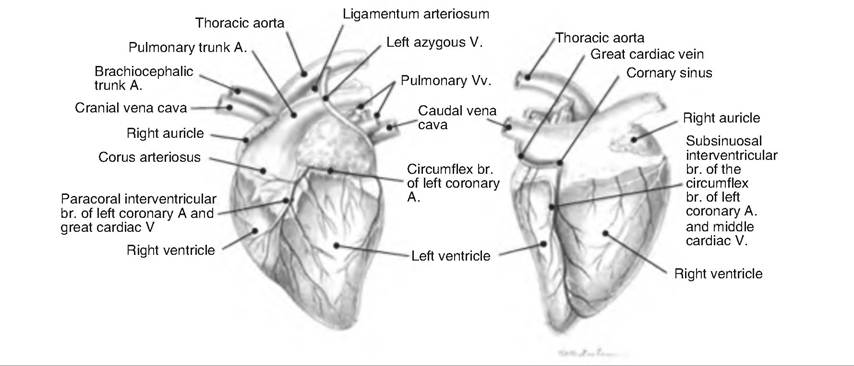
Fig. 13.9. Coronary circulation of goat heart. A., artery; V., vein; br. branch. (Reprinted from Constantinescu, 2001. With permission from the publisher.)
Coronary circulation
The myocardium, or heart muscle, receives nutrients via the coronary circulation (Fig. 13.9). Blood leaves the aorta and passes into the left and right coronary arteries arising at the base of the aorta and encircling the heart in the AV groove. The left coronary artery has two branches. The anterior interventricular artery travels in the anterior interventricular sulcus and supplies the interventricular septum and ventral walls of both ventricles. The circumflex artery supplies the left atrium and dorsal walls of the left ventricle.
The right coronary artery also divides into two branches. The marginal artery supplies the lateral right side of the heart, and the posterior interventricular artery travels to the heart apex and supplies the posterior ventricular walls.
After passing through the capillaries, venous blood in the heart collects in the cardiac veins. These veins carry blood to the coronary sinus, which empties into the right atrium.
Cardiac muscle and the cardiac conduction system
Cardiac muscle
Cardiac muscle is also called involuntary, striated muscle. Cardiac muscle fibers are shorter and less circular than skeletal muscle fibers. They generally contain a single nucleus, although occasionally two are present. Cardiac muscle fibers connect with neighboring fibers via thickening of the Sarcolemma called intercalated discs (Fig. 13.10). These discs contain des- mosomes that hold the fibers together and gap junctions that allow action potentials to move among cardiac muscle fibers. The gap junctions allow the cardiac muscle fibers to act as a functional syncytium, so that the atria and ventricles can contract as a unit.
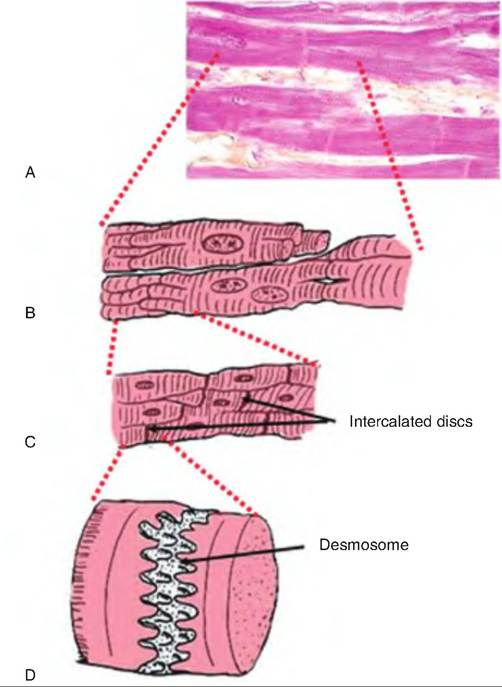
Fig. 13.10. Cardiac muscle (A) Histological section of cardiac muscle. (B) Drawing of a longitudinal section of cardiac muscle showing the branched cardiac muscle fibers. (C) A close-up of the intersection of adjacent cardiac muscle fibers and their intervening intercalated discs. (D) A close-up of the intercalated disc showing a desmosome.
Cardiac muscle fibers contain larger, more numerous mitochondria than skeletal muscle. The mitochondria account for 25% of the cell volume in cardiac muscle while only occupying 2% in skeletal muscle. Like skeletal muscle fibers, cardiac muscle contains sarcomeres. However, the T tubules in cardiac muscle fibers are wider and less abundant than in skeletal muscle. In addition, there is only one T tubule per sarcomere in cardiac fibers, entering at the Z disc, whereas there are two per sarcomere in skeletal muscle fibers entering at the junction of the A and I bands. The sarcoplasmic reticulum (SR) is also less extensive in cardiac muscle fibers.
The conduction system
The heart contains specialized cardiac muscle fibers that can self-generate an action potential, and are therefore called autorhythmic fibers. These cells do not require extrinsic neural input, and they can continue to generate an action potential even when the heart is removed from the body. These autorhythmic cells act as a pacemaker, establishing the basic electrical activity in the heart.
In addition, there are specialized cardiac muscle fibers that form a conduction system that provides a path for electrical excitation to travel throughout the heart (Fig. 13.11). This conduction system helps the heart pump in a coordinated manner so that blood can be pumped throughout the body.
Cardiac electrical activity is propagated through the heart conduction system in the following manner:
1. Located in the wall of the right atria near the entrance of the inferior vena cava is the sinoatrial (SA) node. The cells in the SA node do not maintain a stable resting membrane potential, but instead spontaneously depolarize 75 times ∕min. Since this is faster than in other areas of the heart, the SA node becomes the pacemaker, establishing the sinus rhythm.
2. Depolarization of the SA node results in an action potential that is propagated throughout the atria via the gap junctions between neighboring cardiac muscle fibers.
3. The action potential reaches the AV node, located in the inferior portion of the interatrial septum, above the tricuspid valve. The AV node delays the action potential about 0.1 second before it travels to the ventricles. The delay occurs because the AV fibers are smaller and have fewer gap junctions. This delay allows the atria time to complete their contraction before the ventricles contract.
4. From the AV node, the action potential moves to the AV bundle, also called the bundle of His, located in the superior portion of the interventricular septum. There are no gap junctions between the atria and the ventricles, which are instead insulated from each other by the fibrous skeleton of the heart. This necessitates that the action potential travel through the AV node to reach the ventricles.
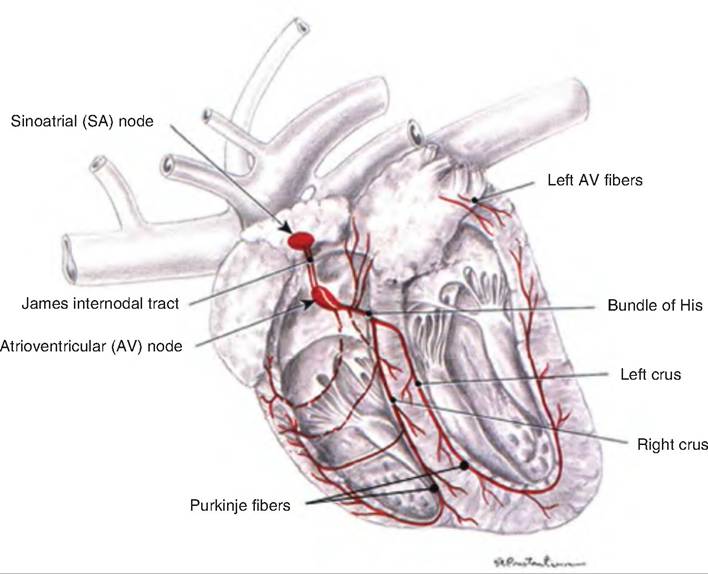
Fig. 13.11. Excitation and conducting system of the dog heart. The pacemaker of the heart is the sinoatrial (SA) node. A wave of depolarization spreads from the SA node throughout the atria. This wave of depolarization then passes through the atrioventricular (AV) node where it is delayed from proceeding to the ventricles. From the AV node, the wave of depolarization travels down the bundle of His located in the intraventricular septum, and then into the Purkinje fibers. (Modified from Constantinescu, 2002.)
5. The action potential continues in the right and left branches. These continue through the inferior portion of the interventricular septum toward the apex of the heart.
6. The right and left branches carry the action potential to the Purkinje fibers, which complete the pathway to the heart apex and then turn superiorly, running up the outer walls of the ventricles toward the atria. Purkinje fibers supply the papillary muscles as well as the ventricular muscles.
The rate at which the SA node depolarizes can be influenced by hormones and the autonomic nervous system. If acetylcholine is released by the parasympathetic nervous system, the SA node slows, while release of epinephrine by the sympathetic nervous system accelerates the SA node.
Mechanisms of heart contraction
Action potentials generated in the heart by the SA node travel throughout the heart via the conduction system as described above. The mechanism of contraction of cardiac muscle fibers is described as follows (Fig. 13.12):
1. Depolarization. As a cardiac muscle fiber is stimulated by a neighboring action potential, voltagegated fast Na+ channels open. This allows a rapid influx of Na+ from the extracellular fluid, which results in depolarization of the cardiac muscle fiber from -90 mV to +30 mV. These channels quickly become inactivated and close.
2. Plateau. As the voltage-gated fast Na+ channels close, voltage-gated slow Ca2+ channels open in the Sarcolemma and SR. The influx of Ca2+ from the extracellular space (20%) causes a large release (80%) of Ca2+ from the SR. Simultaneously, the membrane permeability to K+ decreases. As a result, the membrane remains depolarized at around OmV for about 0.25 second, compared to about 0.001 second in skeletal muscle.
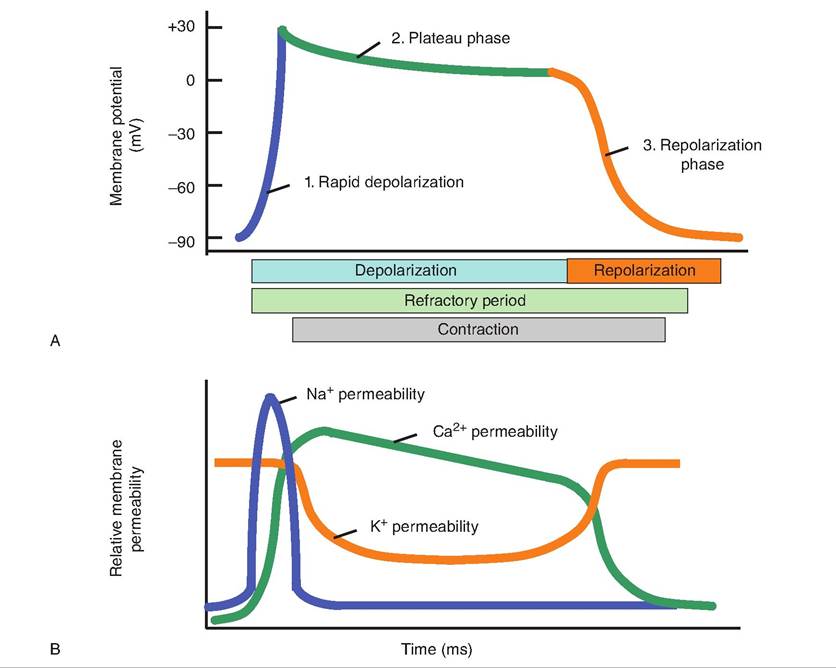
Fig. 13.12. Action potential in cardiac muscle fibers. (A) The action potential in cardiac muscle fibers has a plateau phase not seen in skeletal muscle fibers. (B) The influx of Na+ causes the rapid depolarization phase while increased Ca2+ permeability leads to the plateau phase. Efflux of K+ results in repolarization.
3. Repolarization. After the relatively long plateau phase, voltage-gated K+ channels open, allowing potassium ions to flow out of the cell and the membrane to repolarize. The cell returns to its resting membrane potential of about -90 mV.
4. Refractory period. The refractory period, or time during which the next contraction cannot be triggered, is relatively long in cardiac muscle compared to skeletal muscle. The refractory period prevents cardiac muscle from developing tetanus, and thereby it allows the heart to act as an effective pump rather than developing a sustained contraction.
The mechanism of contraction of cardiac muscle fibers is similar to that in skeletal muscle fibers. As intracellular Ca2+ concentrations increase, Ca2+binds to troponin, causing the tropomyosin to move and thus uncovering the myosin-binding sites on the actin filaments. Myosin then binds to actin, and the actin is pulled across the myosin filament. Drugs that alter the movement of calcium into the cardiac muscle fibers can affect the strength of heart contraction.
ATP production
Cardiac muscle has little capacity for anaerobic cellular respiration; thus, cardiac muscle relies almost entirely on aerobic respiration. Therefore, cardiac muscle needs a continuous supply of O2, which arrives via the coronary circulation or is released from myoglobin inside the cardiac muscle fibers. Cardiac muscle can produce ATP from the oxidation of fatty acids, glucose, lactic acid, amino acids, and ketone bodies.
Cardiac muscle also contains creatine phosphate, which can be used to produce ATP. The enzyme creatine kinase can catalyze the transfer of a phosphate group from creatine phosphate to ADP to produce a new molecule of ATR If the heart is damaged, it releases creatine kinase into the bloodstream, which is often measured as an indicator of heart damage.
Electrocardiogram
The propagation of the action potentials through the heart produces electrical currents that can be detected on the surface of the body. A recording of these electrical activities is called an electrocardiogram (ECG or EKG). An ECG represents all of the electrical activity in the heart rather than a single action potential (Fig. 13.13).
Two electrodes are generally placed on each forelimb and one on the left hind limb. The potential difference between electrodes is measured using different combinations of electrodes. By comparing these various recordings, it is possible to determine whether
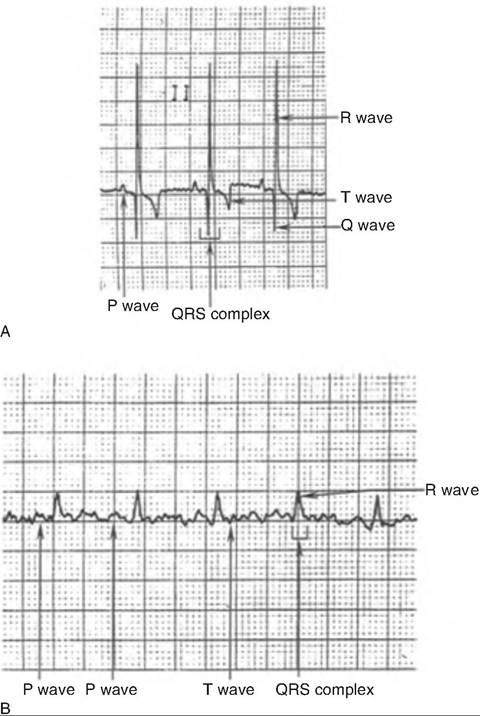
Fig. 13.13. Normal electrocardiogram. (A) Dog. (B) Cat. (Reprinted from Constantinescu, 2002. With permission from the publisher.)
there are abnormalities in the conduction system or whether the heart is damaged.
Each segment of the ECG is generated from a specific area of the heart in sequential manner (Fig. 13.14). A typical ECG has three characteristic waves with each heart beat. The first, or P wave, is a small upward deflection reflecting atrial depolarization (Fig. 13.13). It is generated as the SA node depolarizes, and the action potential spreads throughout the atria. The second wave, or QRS complex, begins with a downward deflection, and then rises sharply and ends with a downward deflection. The QRS complex represents ventricular depolarization. Its shape is complex because the movement of the wave of depolarization through the ventricle changes direction throughout the wave. The third wave is the T wave, and it represents ventricular repolarization. Since repolarization is slower than depolarization, the T wave is longer than the QRS complex.
Within the ECG, it is also possible to examine various intervals or segments. The P-Q interval is the time between the beginning of atrial excitation and the
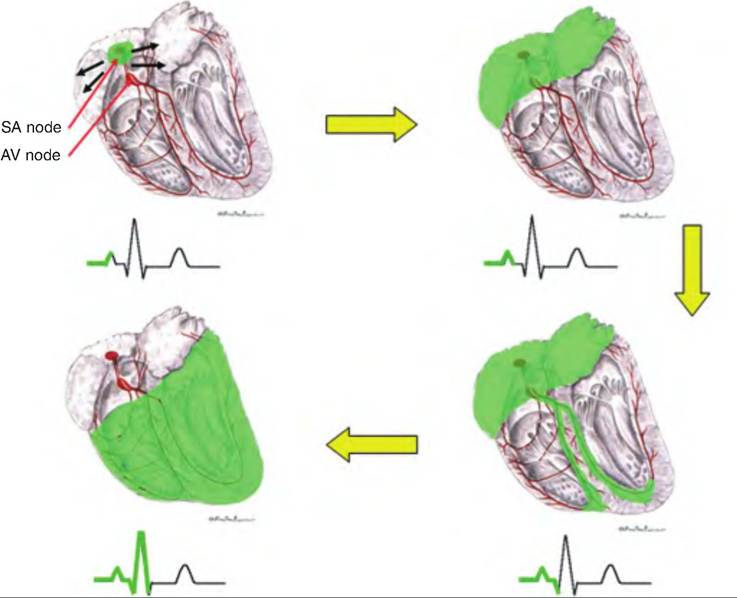
Fig. 13.14. Heart excitation and the ECG tracing. The green shading indicates the relationship between the area of the heart depolarizing coincident with the portion of the ECG produced by that depolarization. (Figure modified from Constantinescu, 2002.)
beginning of ventricular excitation. Therefore, it represents the time for the action potential to travel through the atria, the AV node, and the remainder of the conduction system. This interval can lengthen if there is coronary heart disease or scar tissue in the heart. The S-T segment begins with the S wave and ends with the beginning of the T wave. It represents the time when the ventricle is depolarized during the plateau phase. The T wave can be elevated in acute myocardial infarction. The Q-T interval is the period from the beginning of ventricular depolarization through ventricular repolarization. It can be lengthened by myocardial damage or myocardial ischemia.
Heart sounds
Auscultation involves listening to body sounds, usually with a stethoscope. Four sounds are created during each heart beat, and two of these sounds are clearly audible. These sounds are typically described as 'Tub-dup." The first sound, Iub, is the AV valves closing. This occurs at the beginning of systole as the ventricular pressure increases above the atria pressure, causing the AV valves to close as blood begins returning to the atria. This sound is louder and longer than the second sound. The dup sound is caused by the semilunar valves closing at the beginning of ventricular diastole. The two other sounds, which are less audible, are due to the blood turbulence during ventricular filling and atrial systole.
Heart murmurs include clicking, rushing, or gurgling sounds. Although not always due to a problem, heart murmurs generally indicate a valve disorder. If the valve is stenotic, meaning it has a narrowed opening, a click may be audible when the valve should be fully opened. In contrast, if a swishing sound is heard when the valve should be closed, it may indicate that blood is able to backflow through the valve.
The cardiac cycle
The total of events associated with the movement of blood during one heartbeat is called the cardiac cycle (Fig. 13.15). The contraction and relaxation periods are called systole and diastole, respectively. The following steps describe the cardiac cycle:
1. Mid-to-late diastole. While the heart is relaxed, blood passively returns to the atria and into the ventricles through the opened AV valves. Approximately 70% of ventricular filling occurs during this time.
2. Atrial systole. During atrial systole, the atria contract while the ventricles remain relaxed. Atrial systole begins with depolarization of the SA node, causing an action potential to spread throughout the atria, appearing as the P wave on an ECG. As the atria contract, the remaining 30% of blood is forced through the opened AV valves and into the ventricles. The volume of blood in the ventricles is referred to as the end diastolic volume (EDV).
3. Ventricular systole. While the atria are relaxed (atrial diastole), the ventricles contract, appearing as the QRS complex on an ECG. As the volume in the ventricles decreases, ventricular pressure increases, causing the AV valves to close. For a fraction of a second, all the heart valves are closed, resulting in the Isovolumetric contraction phase. When ventricular pressure exceeds the pressure in the large arteries, the semilunar valves are forced open, leading to the ventricular ejection phase.
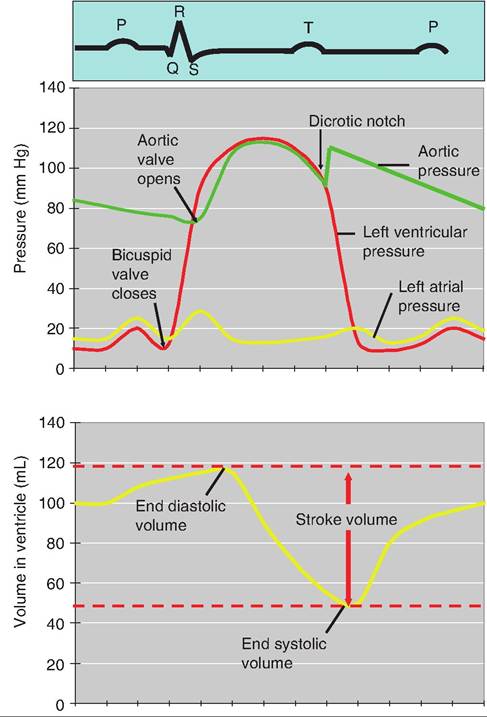
Fig. 13.15. Cardiac cycle. The top of the figure shows a typical ECG. The middle graph displays simultaneous changes in left atrial and ventricle pressure, and aortic pressure. The bottom graph shows changes in ventricular volume during the cardiac cycle.
4. Early diastole. Immediately following the T wave (i.e., ventricle repolarization), the ventricles relax, and the amount of blood remaining in the ventricles is referred to as the end systolic volume (ESV). As the pressure in the ventricles decreases, blood in the aorta and pulmonary arteries begins to return to the heart, causing closure of the semilunar valves. As these valves close, it causes a transient increase in aortic blood pressure (BP) called the dicrotic notch.
If the HR is 75beats/min, the cardiac cycle is about 0.8 second. Atrial systole lasts about 0.1 second, ventricular systole lasts about 0.3 second, and the remainder of the cycle is the quiescent period.
Cardiac output
The amount of blood pumped by the heart can be altered in response to metabolic changes caused by such factors as exercise, environmental temperature changes, or blood loss. The amount of blood pumped by either the right or left ventricle per minute is called the cardiac output (CO). CO is equal to stroke volume (SV), the amount of blood pumped by the ventricle per heartbeat multiplied by the HR:
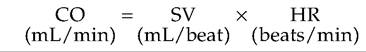
SV is equal to end diastolic volume (EDV) minus end systolic volume (ESV). The heart pumps approximately 60% of the blood in its chambers with each beat.
Factors that can alter SV will alter CO. Cardiac reserve is the difference between an animal's maximum CO and its resting CO (Box 13.7).
Regulation of stroke volume
The heart will pump all the blood returning during systole. Three factors regulate SV: preload, contractility, and afterload.
Preload
Preload is the amount of stretch on the heart prior to contraction. Within limits, greater stretch of the heart results in more forceful contraction. This is known as the Frank-Starling law of the heart. Cardiac muscle fibers are normally shorter than their optimal length for generating force. As a result, stretch of these fibers results in increased contractile force.
The preload is directly proportional to the volume of blood in the ventricles, or EDV Two factors affect EDV: (1) duration Ofventricular diastole and (2) venous return, the amount of blood returning to the heart. As HR increases, duration of diastole shortens, resulting
Box 13.7 Canine heartworms
Canine heartworms, Dirofilaria immitus, are common in the hearts and major heart blood vesicles of dogs throughout the world. The male worms are a few inches in length; the female worms are about double the size and cause most of the damage. The worms are transferred from dog to dog through the bite of an infected mosquito. Mosquitoes transfer microscopic larva that migrate through the body and arrive at the heart several months later where they mature into adult worms. Damage to the dog's heart is due to adult worms.
The first sign of heartworm disease is often premature aging in which dogs gray prematurely around the muzzle and forelegs. Then their activity level decreases and their coats lack luster. Further progression results in a chronic dry cough most noticeable at night when the dog is resting or in a sitting position. At the same time, the dog's heart and pulmonary arteries enlarge due to mechanical obstruction of the worms, inflammation, and valvular damage to the heart.
Until recently, the only medicine available to cure infected dogs of heartworms contained arsenic. More recent medications have an added ingredient, pyrantel pamoate, which prevents infestation with hookworms and roundworms as well.
in a smaller EDV and a smaller SV. Although the decreased SV can be offset by the increased HR, if HR becomes too rapid, there is insufficient preload and CO declines. In contrast, during exercise, venous return increases because of increased squeezing of skeletal muscle on the veins. Consequently, SV increases.
Contractility
Contractility is the strength of contraction at a given preload, and it is independent of muscle stretch and EDV Although preload is the major intrinsic factor regulating SV, contractility is influenced by extrinsic factors. Substances that increase contractility are called positive inotropic agents while those that decrease contractility are called negative inotropic agents.
Positive inotropic agents generally stimulate Ca2+ influx into the cytosol of cardiac muscle fibers, strengthening the force of contraction. Such agents include digitalis, glucagon, thyroxine, norepinephrine (NE), and epinephrine (Fig. 13.16). Negative inotropic agents, which impair Ca2+ inflow, include anoxia, acidosis, increased extracellular K+ levels, and calcium channel blockers.
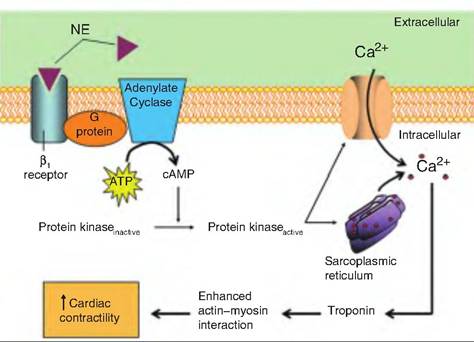
Fig. 13.16. Mechanism of norepinephrine (NE)-induced increase in heart contractility. NE, or epinephrine, binds to β1- adrenoreceptor on the cardiac muscle fibers. This activates a G protein, which activates adenylate cyclase, thus producing cAMP. cAMP then activates protein kinase, which causes increased cytosolic Ca2+, coming both from the extracellular space and the sarcoplasmic reticulum. Ca2+ increases contractility.
Afterload
The pressure that must be exceeded by the ventricles before blood can be ejected through the semilunar valves is called afterload. Any factor that increases afterload will increase ESV and decrease SV Such factors include hypertension or narrowing of the arteries, as in arteriosclerosis.
Regulation of heart rate
Cardiac output depends on heart rate (HR) and SV Changes in HR are important in short-term regulation of CO. Factors that increase HR are positive chronotropic factors; those that decrease HR are negative chronotropic factors. The most important factor controlling HR is the autonomic nervous system.
Autonomic nervous system regulation
The cardiovascular center in the medulla oblongata influences HR. This center receives input from sensory receptors, the limbic system, and the cerebral cortex. It directs the output from both the parasympathetic and sympathetic divisions of the autonomic nervous system.
The cardiovascular center receives sensory input from several areas. Proprioceptors monitor the positions of the limbs and joints. Increased movement of joints, such as during exercise, sends signals resulting in a rapid rise in HR. Chemoreceptors monitor blood chemical changes that can lead to changes in HR. Baroreceptors are located in the aortic arch and carotid arteries. Sudden changes in pressure in these regions cause changes in HR.
Activation of the sympathetic nervous system by either emotional or physical factors causes increased HR. This activation occurs via the cardiovascular center, which can stimulate HR via fibers from the spinal cord that stimulate the cardiac accelerator nerves extending from the spinal cord to the SA node, to the AV node, and throughout the myocardium. The sympathetic nerve fibers release NE, which binds to β1 adrenergic receptors in the heart. NE accelerates the rate of depolarization of the SA node and increases Ca2+ influx into cardiac myofibers, increasing contractility. Both of these effects result in enhanced pumping of blood during systole.
Although large increases in HR decrease EDV and SV, moderate increases in HR are associated with increased contractility, which maintains SV, and therefore CO increases.
Activation of the parasympathetic nervous system sends signals to the heart via the vagus nerve (cranial nerve X). The vagus nerve terminates in the SA node, AV node, and atrial myocardium. Parasympathetic fibers release acetylcholine, which decreases the spontaneous rate of depolarization of the SA node. Parasympathetic input has little effect on contractility.
Chemical regulation of heart rate
Chemicals can have a profound effect on the heart. The major chemicals affecting the heart are hormones and cations.
Hormones
Epinephrine and NE are both released from the adrenal medulla, and, acting as neurohormones, increase HR and contractility. Thyroid hormones also increase HR and contractility.
Cations
Extracellular and intracellular cation concentrations are important in maintaining resting membrane potentials. Therefore, it should come as no surprise that alterations in cation concentrations will affect heart function. Elevated blood Na+ concentrations decrease HR and contractility by interfering with Ca2+ influx into the cardiac muscle fiber. Increased blood K+ also decreases HR and contractility, but it does so by inhibiting the formation of the action potential. Increasing blood Ca2+ levels increases HR and contractility by leading to increased intracellular Ca2+.