Blood vessels and hemodynamics
Structure and function of blood vessels
There are five main types of blood vessels: arteries, arterioles, capillaries, venules, and veins. Arteries carry blood away from the heart as they branch or diverge into smaller arterioles that then carry blood to the capillaries.
Blood leaving the capillaries enters venules, which merge into the larger veins that ultimately enter the heart.Blood vessel walls
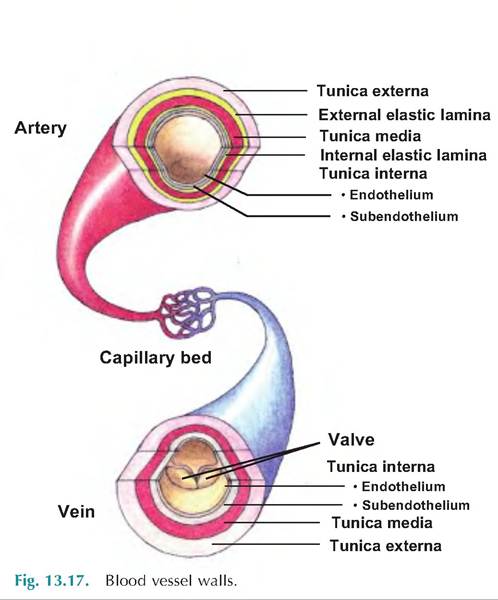
Except in the smallest vessels, there are three layers, or tunics, surrounding the blood vessel lumen (Fig. 13.17). The tunica interna, or tunica intima, is the innermost layer. In intimate contact with the blood, this layer contains the endothelium consisting of simple squamous epithelium lining the lumen. These epithelial cells sit on a loose connective tissue basement membrane called the subendothelial layer. The endothelium is continuous with the endocardium lining the inside of the heart.
The middle layer, or tunica media, consists of a circular layer of smooth muscle and elastin. Stimulation of the vasomotor nerve fibers by the sympathetic nervous system causes vasoconstriction in which the lumen diameter decreases. Relaxation of the smooth muscle results in vasodilation or an increase in lumen diameter.
The outer layer, the tunica externa, or tunica adventitia, is composed of loosely woven collagen fibers. This layer reinforces and protects the vessels, and it is the site where nerve fibers and lymphatic vessels enter to provide nourishment.
Arteries
The arteries near the heart are called elastic arteries because they contain a large proportion of elastic fibers in the tunica media. They are large in diameter, therefore providing little resistance to blood flow. They expand to accommodate blood ejected from the ventricles.
As the blood pressure (BP) decreases, these vessels recoil, thus helping to maintain pressure. They are sometimes called conducting arteries because they carry blood to more muscular, medium-sized vessels. Elastic arteries include the aorta, and the brachiocephalic, common carotid, subclavian, vertebral, pulmonary, and common iliac arteries.The medium-sized arteries are called muscular arteries because they contain more muscle and less elastic fibers in the tunica media. Being more muscular, they have greater capacity to Vasoconstrict. They are sometimes called distributing arteries because they deliver blood to various parts of the body.
Arterioles
The smallest of the arteries, arterioles deliver blood to the capillaries. Large arterioles contain all three tunics, with the tunica media having considerable smooth muscle and few elastic fibers. The smallest arterioles consist of simply a layer of endothelial cells surrounded by scattered smooth muscle cells. A metarte- riole connects an arteriole with 10-100 capillaries making a capillary bed.
Capillaries
Capillaries, also called exchange vessels, are the smallest vessels. Their walls consist of only a tunica interna. Although capillaries are found in most places in the body, they are lacking in epithelium, the cornea and lens of the eye, and cartilage.
True capillaries originate from arterioles or metarte- rioles. At their origin is a ring of smooth muscle called the precapillary sphincter (Fig. 13.18). When contracted, the sphincter restricts the flow of blood into the capillary bed. Normally, blood flow within a capillary bed is intermittent due to changing vasomotor tone in the precapillary sphincter.
There are three types of capillaries: continuous, fenestrated, and sinusoidal (Fig. 13.19). Found in skin and muscles, the most common type is continuous, in which the endothelial cells form an uninterrupted layer with tight junctions between cells. However, there are intercellular clefts, or gaps, between neighboring cells allowing for exchange of nutrients.
Within the brain, continuous capillaries lack intercellular clefts, and therefore form a structural barrier between the blood and brain, called the blood-brain barrier.Fenestrated capillaries are similar to continuous capillaries, but they also have pores, or fenestrations,
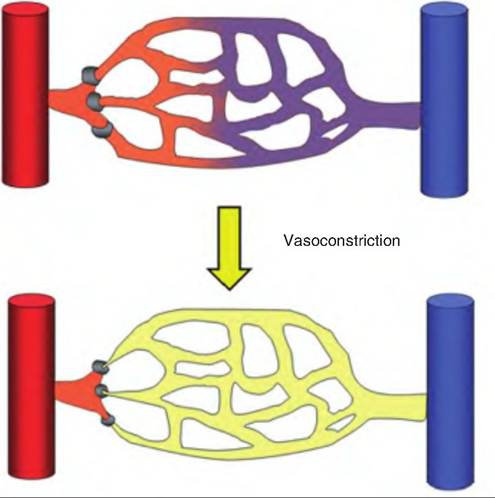
Fig. 13.18. Precapillary sphincters. Constriction of precapillary sphincters restricts the blood flow within a capillary bed.
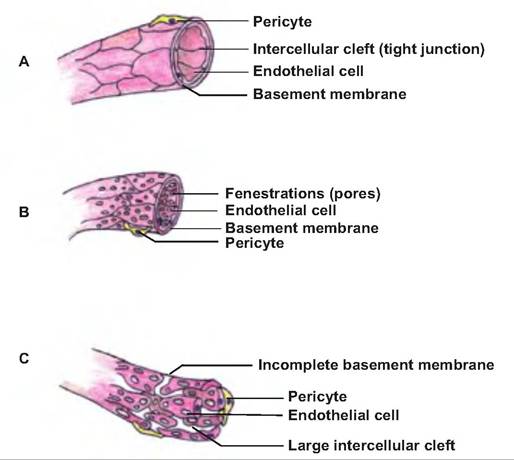
Fig. 13.19. Types of capillaries. (A) Continuous, (B) fenestrated, and (C) sinusoidal.
in the endothelial cells that allow substances to move out of the vessels. Such capillaries are found in the kidneys, villi of the small intestine, choroids plexus, ciliary processes of the eyes, and endocrine glands.
Sinusoidal capillaries have large, irregularly shaped lumens, and their endothelial cells have large fenestrations. They also lack a complete basement membrane, and thus they are very leaky. Such capillaries are found in the liver, bone marrow, lymphoid tissue, such as the spleen, anterior pituitary, and parathyroid glands.
Venules
The smallest venules found close to capillaries consist of a tunica interna and a tunica media with a few smooth muscle cells. As the venules enlarge, they may contain a tunica externa.
Veins
Veins have the same three layers as arteries, but their thicknesses vary. The tunica interna and tunica media are thinner. The tunica externa is the thickest layer, containing collagen and elastic fibers. In the largest veins, the tunica externa also contains a longitudinal smooth muscle layer.
Unlike other vessels, veins also contain venous valves (Fig. 13.20). Formed from the tunica interna, these one-way valves point toward the heart. The contraction of skeletal muscle during movement and increased thoracic pressure associated with respiration squeezes the veins, forcing blood toward the heart.
As the skeletal muscle relaxes, the backflow of blood is prevented by the venous valves.Because veins have a large lumen and thin walls, they can contain a large blood volume. Because veins can contain up to 65% of blood volume, they are called capacitance vessels.
Anastomoses
Most tissues receive blood from multiple arteries. The merging of these multiple sources results in an anastomosis. Such mergers provide alternate routes by which tissue can receive blood. If blood flow through one artery is prevented by an occlusion or loss of a
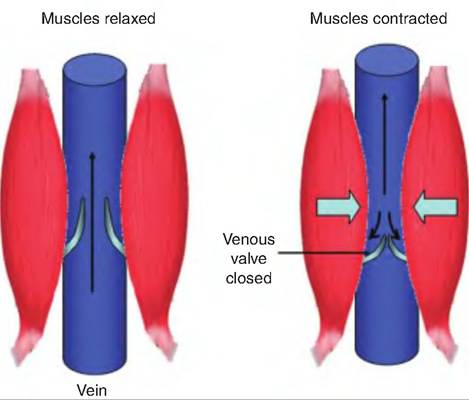
Fig. 13.20. Venous blood flow. Contraction of skeletal muscle and increased thoracic pressure due to respiration squeezes the veins, which causes the blood to move toward the heart. The blood is prevented from flowing backward, away from the heart, by one-way venous valves located in the veins.
vessel, blood still goes to the tissue through an anastomosis, thus providing collateral circulation.
Portal systems
In several places throughout the body, there are also vessels that link one capillary bed to another. These are known as portal vessels. Such vessels have the histological structure of veins. The complex of two capillary beds and the intervening portal vessel is known as a portal system.
One such portal system, called the hypophyseal portal system, consists of a capillary network in the median eminence supplied by the superior hypophyseal artery. This capillary network unites to form a series of vessels that spiral around the infundibulum and carry blood to a second capillary network located in the anterior pituitary.
A second example of a portal system is the hepatic portal system. The capillaries along the digestive system deliver blood into the inferior mesenteric vein, the splenic vein, and the superior mesenteric vein. These all deliver blood to the hepatic portal vein, which forms from a fusion of the superior mesenteric and splenic veins.
The hepatic portal vein then carries blood to a capillary bed located in the liver. While carrying blood to the liver, the hepatic portal vein receives more blood from the gastric veins draining the stomach and the cystic vein coming from the gallbladder.Capillary exchange
The purpose of the circulatory system is to deliver nutrients and remove wastes from tissues. This occurs through capillary exchange, in which substances move between the blood and interstitial fluid.
Diffusion
Capillary exchange generally occurs by simple diffusion, where chemicals move along their concentration gradient moving from an area of higher concentration to an area of lower concentration. O2 and nutrients are generally in higher concentrations within the blood and therefore pass into the tissue; waste products are in higher concentration in the interstitial space and pass into the blood.
In all capillaries except sinusoids, the space between endothelial cells prevents the movement of plasma proteins from leaving the capillaries. In contrast, water-soluble chemicals, including glucose and amino acids, pass out of the capillaries through fenestrations of intercellular clefts. Lipid-soluble materials such as O2, CO2, and steroid hormones pass directly through the endothelial cell wall. In liver sinusoids, the gaps
between the endothelial cells are large enough to allow proteins synthesized in the liver to enter the blood.
Bulk flow
The passive movement of large numbers of materials across a membrane is called bulk flow. Bulk flow moves from an area of higher pressure to an area of lower pressure. Diffusion accounts for most nutrient exchange across the capillary wall; bulk flow controls blood and interstitial fluid volume. The movement of fluid and solutes from capillaries into the interstitial space is called filtration, and the movement from interstitial fluid into the capillaries is called reabsorption.
Capillary hydrostatic pressure (HPc) is the force exerted by blood against the capillary wall.
This pressure tends to force fluid out of the capillary at the arteriole end of a capillary bed. HPc is opposed by the interstitial fluid hydrostatic pressure (HPif), which pushes inward against the capillary. Therefore, the net hydrostatic pressure (Net HP) acting on a capillary is HPc minus HPif. However, HPif is generally zero, so the effective hydrostatic pressure acting on a capillary is equal to HPc. HPc is larger at the arterial end of a capillary than at the venule end of the capillary.HPc is also opposed by the colloid osmotic pressure (OPc), or oncotic pressure. This is the osmotic pressure inside the capillary caused by the presence of large plasma proteins that cannot leave the capillaries. The main protein responsible for OPc is albumin. OPc does not vary between the arterial and venule end of the capillary. Since the interstitial fluid has a few proteins, there is also an interstitial fluid osmotic pressure (OPif) opposing OPc. OPif is generally only around 1 mmHg.
Net filtration pressure (NFP), which is an interaction of hydrostatic and osmotic pressures, determines the direction of movement of fluids across the capillary wall (Fig. 13.21). NFP is calculated as follows:
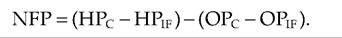
If we assume that at the arterial end of a capillary HPc is 40 mm Hg, OPc is 25 mm Hg, and OPif is 1 mmHg, then:
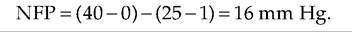
If, at the venule end of the capillary, HPc is 20 mm Hg, OPc is 25 mm Hg, and OPif is ImmHg, then:
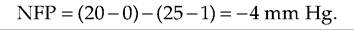
Since HPc varies along the capillary, there is a net movement of materials out of the capillary at the arterial end of the capillary and a net movement inward at the venule end. About 85% of the fluid filtered at the capillary is reabsorbed. The remaining fluid enters the lymphatic capillaries and eventually returns to the circulation at the subclavian vein.
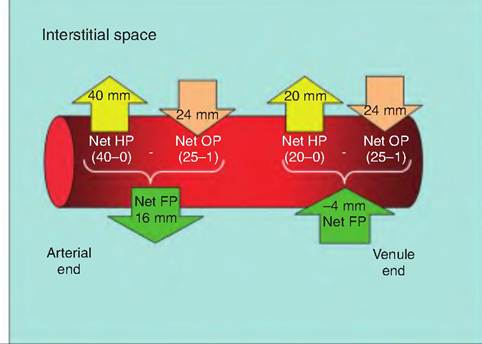
Fig. 13.21. Fl uid flow across the capillary wall. At the arteriole end of a capillary, the hydrostatic pressure inside the capillary exceeds the oncotic pressure inside the capillary, which results in a net outward filtration pressure. In contrast, at the venule end of the capillary, the oncotic pressure inside the capillary exceeds the outwardly directed hydrostatic pressure, causing a net inwardly directed filtration pressure. The interstitial fluid that does not return to the capillary enters the lymphatic system.
Lack of reabsorption or an increase in filtration leads to edema, an abnormal increase in interstitial fluid volume. Lack of reabsorption can be caused by a decreased concentration of plasma proteins as is seen during liver disease, burns, malnutrition, or kidney disease. An increase in filtration can result from increased capillary pressure or damage to the endothelial wall caused by chemical, mechanical, or bacterial agents.
Factors affecting blood flow
Flow, pressure, and resistance
Blood flow (F) refers to the volume of blood flowing through a tissue during a given period of time. Total blood flow is equal to CO. Blood flow is directly proportional to the difference in BP between two points, and is inversely proportional to the resistance (R) to blood flow in the vessels:
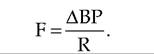
Since blood vessels have a great capacity to vaso- Constrict and vasodilate, R has a greater affect on F than does BP Since total blood flow is equal to CO, CO equals ΔBP∕R.
Blood pressure
Blood flow occurs because of the pumping action of the heart. BP is the hydrostatic pressure exerted by the
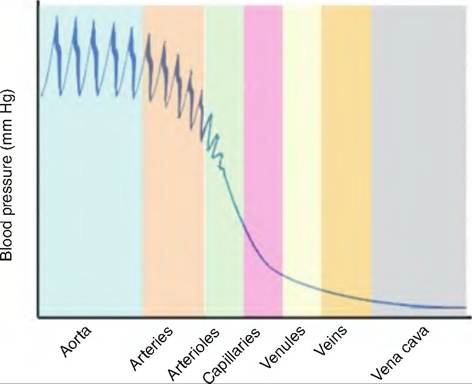
Fig. 13.22. Blood pressure throughout the circulatory system. The pulse pressure in the large arteries is substantial. As the blood moves through the circulatory system, pulse pressure declines. Although capillaries have a small cross-sectional area, blood pressure drops in the capillary bed due to the large numbers of capillaries. Blood pressure within the veins is low. (Modified from Marieb, 2004.)
blood against the blood vessel wall. BP occurs because there is resistance to blood flow. BP is highest in the arteries and decreases as blood moves through the circulatory system (Fig. 13.22).
BP is highest at the end of ventricular contraction, which is called systolic blood pressure. During diastole, the semilunar valves close, preventing blood from returning to the heart, and the elastic recoil in the large arteries maintains pressure that keeps blood flowing. At the end of diastole, the BP is at its lowest point, which is called diastolic blood pressure.
Pulse pressure is the difference between systolic and diastolic blood pressure. Mean arterial pressure (MAP) is the pressure that propels blood. Since the heart spends more time in diastole than systole, MAP is not simply the mean of diastolic and systolic blood pressure. Instead, MAP is equal to diastolic blood pressure plus one-third of pulse pressure:

Resistance
Vascular resistance opposes blood flow, and is due to the friction between blood and the vessel walls. As shown in Figure 13.22, MAP and pulse pressure decline with increasing distance from the heart. This is due to the increased friction between the blood and vessel walls, and the decreasing elasticity of the vessel walls farther from the heart.
Vascular resistance is dependent on the size of the blood vessel lumen, blood viscosity, and total blood vessel length.
Blood vessel lumen
As lumen size decreases, resistance to blood flow increases. Resistance is inversely proportional to the fourth power of the lumen diameter. Therefore,
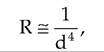
where d = lumen diameter. A small change in blood vessel diameter results in a large change in resistance. Decreasing the diameter of a vessel by one-half will increase resistance by 24, or 16-fold. Therefore, vasodilation and vasoconstriction have a large affect on vascular resistance.
Blood viscosity
Blood viscosity is affected by the concentration of erythrocytes. Increasing the erythrocyte concentration, that is, increasing the hematocrit, increases blood viscosity. This can be a result of dehydration or polycythemia. In contrast, decreased viscosity can result from hemorrhage or anemia.
Total blood vessel length
Resistance to blood flow is proportional to the blood vessel length. The longer the blood vessel, the greater the resistance. Hence, obesity in animals can result in hypertension due to increased length of blood vessels associated with adipose tissue.
Venous blood return
Blood pressure within the veins is relatively low due to the cumulative effects of peripheral resistance throughout the vascular system (Fig. 13.21). Therefore, there are other factors besides the heart that are important in venous circulation. First, the respiratory pump involves increases in abdominal pressure associated with inhalation. This increase in pressure squeezes venous blood toward the heart. As abdominal pressure is increasing, thoracic pressure is decreasing during inhalation, further allowing blood to enter the right atrium.
Second, there is a muscular pump that aids in venous return. As an animal moves, the skeletal muscle squeezes the veins, thus moving the blood toward the heart. As the muscles relax, the one-way valves in the veins prevent the backflow of blood.
Maintaining blood pressure
There are both short- and long-term mechanisms controlling BP and blood flow. These mechanisms are responsible for controlling HR, SV, systemic vascular resistance, and blood volume.
Neural regulation
The cardiovascular center is located in the medulla oblongata and is responsible for controlling HR and SV. As discussed above, the cardiovascular center sends sympathetic signals via the cardiac accelerator nerve that increases HR and contractility. Conversely, parasympathetic signals from the cardiovascular center are carried via the vagus nerve and decrease HR and contractility. The cardiovascular center also sends signals to blood vessels via the vasomotor nerves, resulting in vasomotor tone, or a moderate amount of constriction within the vessels. The vasomotor tone can be altered by either vasoconstriction or vasodilation.
The cardiovascular center is involved in two reflexes controlling BP: the baroreceptor reflex and chemoreceptor reflex.
Baroreceptor reflex
There are pressure-sensitive mechanoreceptors located in swellings within the internal carotid arteries, known as the carotid sinuses, the aortic arch, and the walls of most large arteries in the neck and thorax. In response to stretch, these receptors send signals resulting in inhibition of the cardiovascular center. This results in vasodilation and a decrease in BP. Conversely, a sudden decrease in BP results in stimulation of the cardiovascular center, a resulting vasoconstriction, and an increase in BR
This reflex is important in making rapid adjustments in BP in response to acute changes. For example, as an animal stands, the pressure in the neck may decrease. The baroreceptor reflex quickly increases BP to maintain adequate flow to the brain.
Chemoreceptor reflex
Chemoreceptors located in the carotid bodies and aortic bodies monitor blood O2, CO2, and H+ concentrations. Hypoxia, acidosis, or hypercapnia (i.e., increased blood CO2) stimulate the chemoreceptors. The resulting signals stimulate sympathetic output from the cardiovascular center, causing vasoconstriction and increased BP
Chemical regulation of blood pressure:
Short-term control
There are several hormones and neurotransmitters that have significant affects on BR Some act directly on blood vessels, and others have differential affects on organs.
Renin-angiotensin-aldosterone (RAA) system
A decrease in BP or blood flow to the kidneys causes the juxtaglomerular cells of the kidneys to secrete
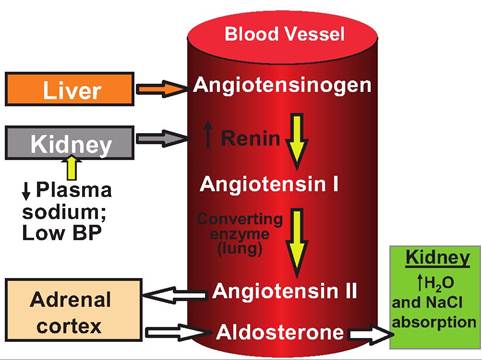
Fig. 13.23. Production and action of renin. Renin is produced by the juxtaglomerular cells in the kidney in response to low blood pressure. Renin converts angiotensinogen, produced by the liver, to angiotensin I, which is then converted to angiotensin Il (ANG II) by a converting enzyme located in the lungs. ANG Il acts on the adrenal cortex to cause the release of aldosterone, causing the kidneys to reabsorb Na+, thus raising blood volume and blood pressure. ANG Il also acts directly on blood vessels causing vasoconstriction.
renin. Renin acts on angiotensinogen produced in the liver to produce angiotensin I (ANG I). As ANG I travels through the lungs, it is converted to angiotensin II (ANG II), which raises BP two ways (Fig. 13.23). First, it causes vasoconstriction, which increases vascular resistance, thereby increasing BR Second, it causes the release of aldosterone from the adrenal cortex. Aldosterone increases sodium and water reabsorption by the kidneys. This results in an increase in blood volume and thereby raises BP
Epinephrine and norepinephrine
Sympathetic stimulation causes the adrenal medulla to secrete both NE and epinephrine. These neurohormones increase the rate and force of heart contractions, thereby increasing CO. They also have a differential effect on various vascular beds, causing vasoconstriction in the skin and visceral organs, while causing vasodilation in skeletal muscle.
Antidiuretic hormone (ADH)
ADH is secreted from the posterior pituitary. It acts in the kidneys to increase water reabsorption, and it also acts directly on blood vessels to cause vasoconstriction. Both these effects cause an increase in BR
Atrial naturiuretic peptide (ANP)
ANP is released from the atria of the heart in response to high BP, that is, stretching of the atria. This hormone decreases sodium reabsorption from the kidneys,
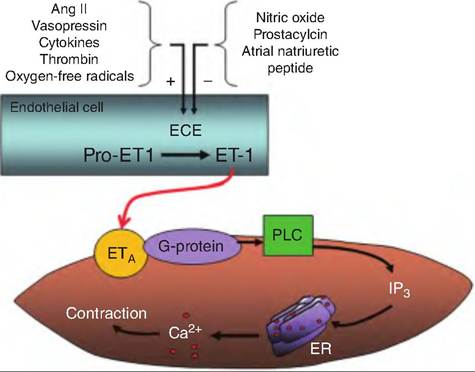
Fig. 13.24. Endothelin production and action. Endothelin is produced by endothelial cells lining the blood vessels. Various stimuli activate endothelin-converting enzyme (ECE), which converts Pro-endothelin-1 (Pro-ETI) to endothelin-1 (ET-1). ET-I diffuses to vascular smooth muscle cells and binds to the ETa receptors. These receptors are coupled to a G protein that activates phospholipase C (PLC), which results in the production of inositol triphosphate (IP3). IP3 causes the release of ETa Ca2+ from the endoplasmic reticulum (ER), which leads to smooth muscle contraction.
thereby decreasing water reabsorption from the kidneys. This results in decreased blood volume and BP.
Endothelial-derived factors
Several chemicals affecting vasomotor tone are produced from the endothelial lining of blood vessels. Endothelin release is stimulated by angiotensin II, ADH, thrombin, cytokines, reactive oxygen species, and shearing forces acting on the vascular endothelium. Its release is inhibited by NO, prostacyclin, and ANP. Endothelin causes vasoconstriction (Fig. 13.24).
Endothelial cells also release a potent vasodilator called NO. Originally called endothelial-derived relaxation factor, NO is a gas produced in response to high BP, acetylcholine, and bradykinin. It diffuses to the neighboring smooth muscle cells where it causes the production of cGMP resulting in relaxation and vasodilation.
Inflammatory chemicals
Erythemia, or vasodilation, is associated with inflammation. A number of chemicals are involved in this response, including histamine, prostacyclin, and kinins. This response allows monocytes and neutrophils to leave the bloodstream and move toward the site of inflammation.
Renal regulation of blood pressure: long-term control
Although most short-term BP control mechanisms work by altering peripheral resistance, long-term renal control alters blood volume (Fig. 13.25). Blood volume has a direct effect on CO, and therefore affects BP An increase in EDV increases CO, thus increasing BP
Direct renal mechanism
An increase in BP or blood volume causes an increased filtration rate in the kidney. As the filtration rate exceeds the kidney tubules reabsorption rate, more urine is produced, resulting in increased fluid loss and decreased blood volume.
Indirect renal mechanism
If arterial BP declines, the kidneys release the enzyme renin into the bloodstream. As shown in Figure 13.23, renin induces angiotensin II (ANG II) production. ANG II has three effects that increase BR First, it causes vasoconstriction. Second, it stimulates the adrenal cortex to secrete aldosterone, a hormone that stimulates sodium reabsorption from the kidney tubules. Third, ANG II stimulates ADH release from the posterior pituitary. ADH increases water reabsorption from the kidney tubules. Increased sodium and water reabsorption from the kidneys leads to increased blood volume and BR
Autoregulation of blood pressure
Local oxygen needs change throughout the body. In order to accommodate these local needs, capillary beds can alter their vasomotor tone in response to local physical and chemical factors.
Physical factors
Increased local temperature, such as occurs during inflammation or exercise, causes vasodilation. Decreased local temperature causes vasoconstriction. In addition, the smooth muscles in arteriole walls display myogenic responses, in which increased stretch causes enhanced contraction while decreased stretch caused relaxation. Such responses help regulate local blood flow as follows: If blood flow through an arteriole decreases, the arteriole wall is stretched less, resulting in smooth muscle relaxation and vasodilation. The vasodilation increases blood flow through the arteriole.
Chemical factors
Cells such as platelets, blood cells, smooth muscle, macrophages, and endothelial cells release a variety of chemicals that modify blood vessel diameter. Metaboli- cally derived vasodilators include K+, H+, lactic acid, and ATP Other tissue-synthesized vasodilators include NO produced by endothelial cells, histamine from mast cells, and monocytes and kinins produced during inflammation. Vasoconstrictors include thromboxane
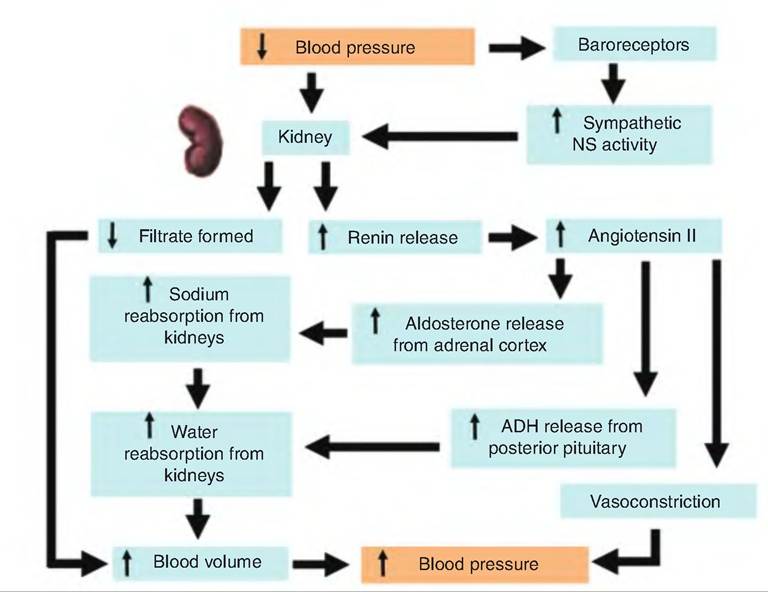
Fig. 13.25. Renal control of blood pressure.
A2 and serotonin from platelets, superoxide radicals, and endothelins from endothelial cells.
Note that the systemic and pulmonary circulatory systems respond differently to changes in O2 levels. Systemic blood vessels dilate in response to low O2, whereas pulmonary blood vessels constrict. Therefore, systemic vessels dilate in order to deliver more O2 to needed areas while pulmonary vessels constrict so that blood is diverted from poorly ventilated alveoli.
Shock and homeostasis
Circulatory shock includes any condition in which blood vessels are unable to deliver adequate O2 and nutrients to meet cellular needs. This results in hypoxia in the affected tissue, leading to a switch to anaerobic metabolism, lactic acid accumulation, and possibly tissue death.
Types of shock
The most common type of shock is hypovolemic shock resulting from massive blood loss. Blood loss can occur internally from the rupture of an artery, or externally from trauma. Excessive loss of body fluids such as occurs in profuse sweating, diarrhea, or vomiting can also cause hypovolemic shock. Treatment involves the restoration of fluids.
Cardiogenic shock occurs when the heart fails to adequately pump. This can be caused by a myocardial infarction, ischemia of the heart, heart valve problems, impaired contractility of the heart, or arrhythmias.
Vascular shock is a result of abnormal expansion of the vascular bed. Although there is no change in blood volume, there is a drastic drop in peripheral resistance leading to a drop in BP. Causes can include anaphylactic shock from an allergic reaction or neurogenic shock resulting from head trauma damaging the cardiovascular center. Another cause can be septic shock resulting from bacterial toxins.
Circulatory routes
Pulmonary circulation
The pulmonary circulation functions to carry deoxygenated blood to the alveoli (air sacs) in the lungs where gas exchange occurs (Fig. 13.26). The blood picks up oxygen in exchange for carbon dioxide. The blood is pumped from the right ventricle into the pulmonary trunk, which then divides into the right and left pulmonary arteries. The pulmonary arteries divide into lobar arteries following the bronchi into the lungs. They then branch, eventually forming pulmonary capillaries in the air sacs. These capillary beds drain into the pulmonary veins. The pulmonary veins return to
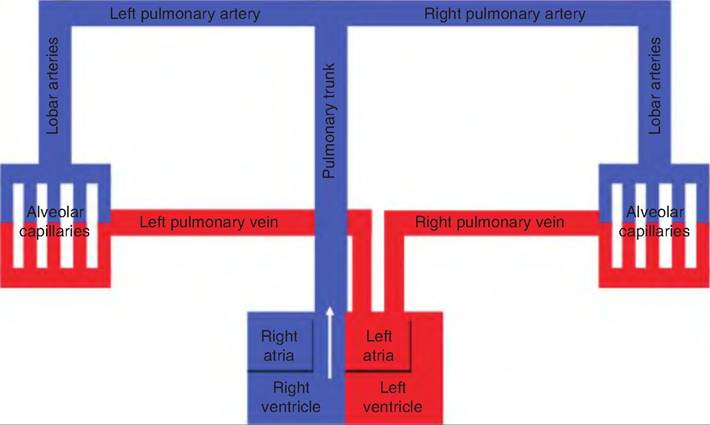
Fig. 13.26. Schematic diagram of pulmonary circulation. The pulmonary circulation carries deoxygenated blood to the lungs where the blood picks up oxygen in exchange for carbon dioxide. The blood returns to the left side of the heart where it enters the systemic circulation.
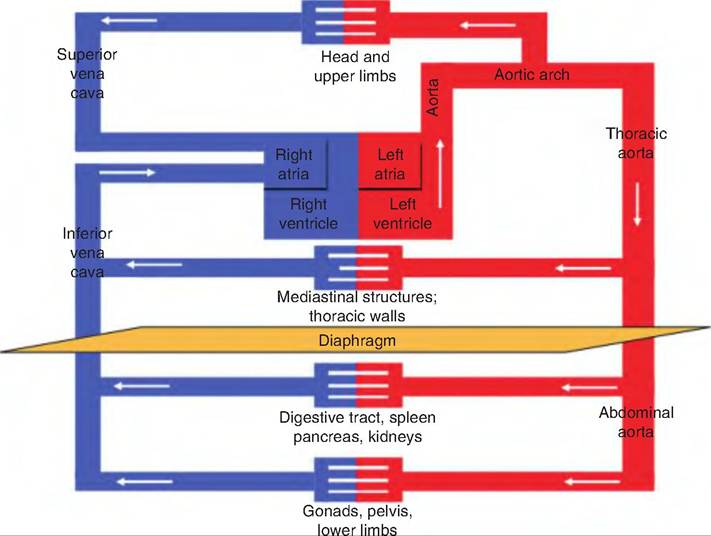
Fig. 13.27. Schematic diagram of systemic circulation. Blood delivered to the left side of the heart from the pulmonary circulation (pulmonary circulation has been omitted from this figure) leaves via the aorta to be circulated throughout the body. After passing through the capillary beds, the blood returns to the right side of the heart via the superior or inferior vena cava.
the left atria from which the blood enters the left ventricle and is then delivered to the body.
Systemic circulation
The systemic circulation includes vessels that deliver oxygenated blood from the left ventricle, throughout the body, and returns deoxygenated blood to the right atrium (Fig. 13.27). The systemic circulatory system includes vessels supplying the tissue needs of the lungs, as well as all the remaining tissues in the body. So, while the pulmonary circulatory system supplies blood only to the gas exchange portion of the lungs, the systemic circulation must supply blood to every
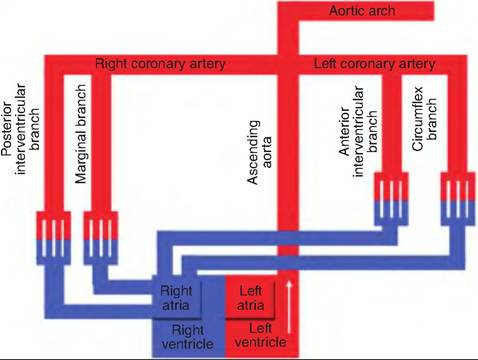
Fig. 13.28. Ascending aorta. The ascending aorta begins at the aortic valve and ascends dorsal to the aortic arch. It gives rise to vessels supplying the heart.
tissue of the body. This is why the left ventricle has a thicker muscular wall than the right ventricle.
Systemic circulation begins with blood traveling through the aorta, and ends with the blood returning via the superior vena cava, inferior vena cava, or coronary sinus.
The aorta has four major divisions: the ascending aorta, arch of the aorta, thoracic aorta, and abdominal aorta. The ascending aorta emerges from the left ventricle and runs posterior to the pulmonary trunk. It gives rise to two coronary arteries that supply the myocardium (Fig. 13.28). It then curves left, giving rise to the aortic arch. It then continues caudally running close to the vertebral bodies. As it courses caudally, it is called the thoracic aorta until it passes through the diaphragm, becoming the abdominal aorta.
The aortic arch is a continuation of the ascending aorta. It gives rise to three major arteries: the brachiocephalic trunk, the left common carotid, and the left subclavian (Fig. 13.29). The brachiocephalic trunk gives rise to the right subclavian artery and right common carotid artery. These vessels provide the arterial supply to the head, neck, front limbs, and a portion of the thoracic spine (Fig. 13.30).
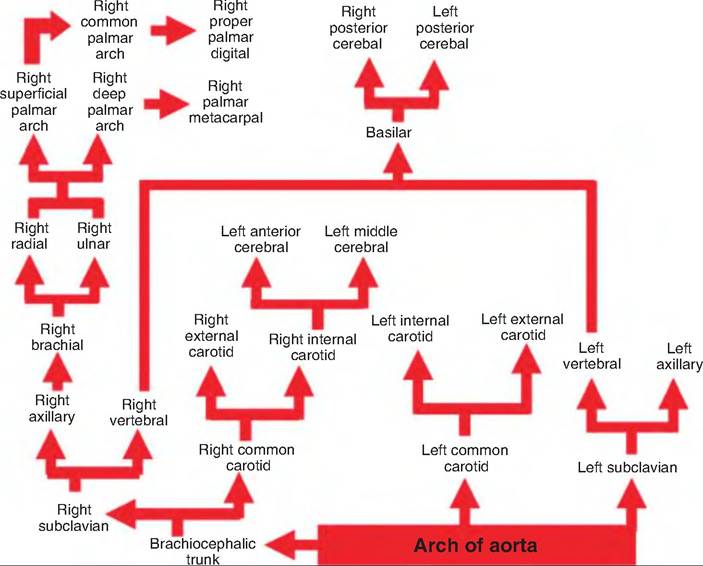
Fig. 13.29. Aortic arch. The aortic arch is a continuation of the ascending aorta.
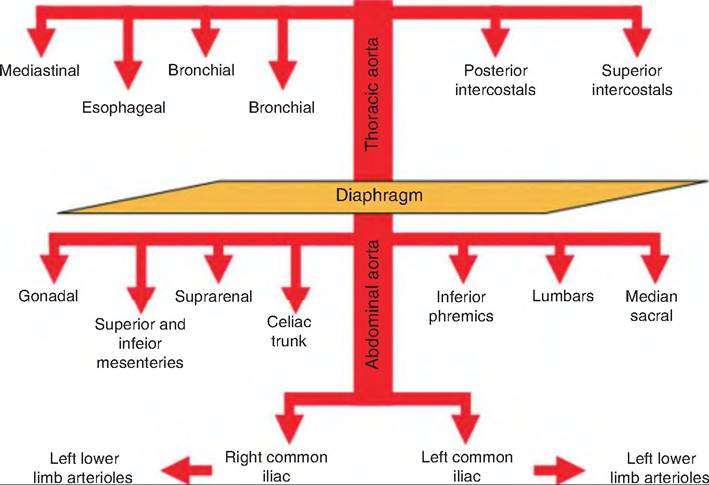
Fig. 13.30. Thoracic and abdominal aorta. The thoracic aorta gives rise to visceral and parietal branches supplying the thorax wall and viscera. The abdominal aorta supplies the abdominal walls and viscera. It ends in the right and left common iliac arteries supplying the pelvis and hind limbs.