Transport of oxygen and carbon dioxide
Hemoglobin and oxygen transport
Oxygen has a low solubility coefficient, so it does not readily dissolve in blood. Instead, over 98% of O2 is bound to hemoglobin. The heme portion of hemoglobin contains four atoms of iron, each able to bind to one molecule of O2.
Adult hemoglobin consists of two alpha and two beta chains. Fetal hemoglobin contains two alpha and two gamma chains. More than 400 different types of abnormal hemoglobin have been identified, but the most common associated with diseases in humans are the following:
• Hemoglobin S. This type of hemoglobin is present in sickle cell anemia.
• Hemoglobin C. This is another type of hemoglobin found in sickle cell anemia.
• Hemoglobin E. This type of hemoglobin is found in people of Southeast Asian descent.
• Hemoglobin D. This type of hemoglobin may be present with sickle cell anemia or thalassemia.
• Hemoglobin H (heavy hemoglobin). This type of hemoglobin may be present in certain types of thalassemia (Box 14.4).
Oxygen bound to hemoglobin forms oxyhemoglobin. Hemoglobin that has released O2 is called reduced hemoglobin, or deoxyhemoglobin. Because it picks up a H+ ion after releasing O2, reduced hemoglobin is abbreviated as HHb:
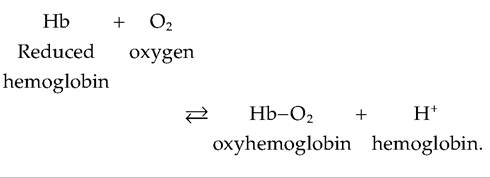
Box 14.4 Onion toxicity in cats and dogs
Allium species (meadow garlic, nodding onion, Pacific onion, and wild garlic) contain Organosulfur compounds that can be very toxic when consumed by cats or dogs. These compounds can cause oxidative hemolysis, which results in cell lysis of erythrocytes. Consuming as little as 5g∕kg or 15-30 g/kg of onions in cats and dogs, respectively, can cause this problem. Treatment includes inducing emesis in asymptomatic dogs and cats known to have ingested these products.
If severely affected, a blood transfusion and supplemental oxygen may be necessary.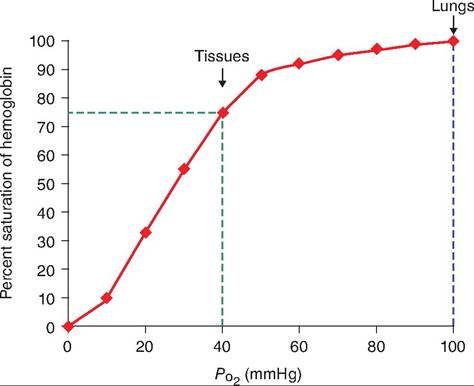
Fig. 14.9. Oxygen-Iiemoglobin dissociation curve. The saturation of hemoglobin is affected by changes in Pθ2. At the lungs, hemoglobin is 100% saturated. As the blood passes through tissue in which the Pθ2 is lower, hemoglobin is able to rapidly unload O2
Pθ2 is the most important factor controlling how much O2 is bound to hemoglobin. When reduced hemoglobin is converted to oxyhemoglobin, it is fully saturated. When only a portion of hemoglobin exists as oxyhemoglobin, it is partially saturated. This can be graphically represented as an oxygen-hemoglobin dissociation curve (Fig. 14.9). At the level of the lungs, hemoglobin becomes fully saturated. As blood passes through tissue where the Pq2 drops to 40 mmHg, hemoglobin unloads its oxygen and is only 75% saturated.
The oxygen-hemoglobin dissociation curve has a steep slope between 10 and 50 mmHg Pθ2. When an animal is at rest, hemoglobin releases approximately only 25% of its oxygen. It maintains a reserve that is available when needed. If the animal begins vigorous exercise, hemoglobin is able to respond by releasing a greater amount of O2.
Other factors affecting the oxygen- hemoglobin dissociation curve
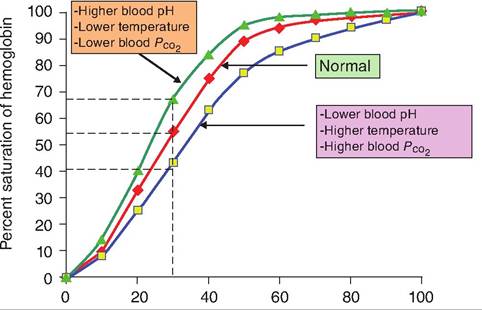
Although the Pj2 is the most important factor affecting hemoglobin's affinity for oxygen, other factors can also influence this association. These factors can shift the oxygen-hemoglobin curve to either side of normal (Fig. 14.10):
1. pH of the blood. Increasing the acidity of blood, that is, lowering the pH, lowers the affinity of hemoglobin for O2. Therefore, as the metabolism of tissue increases, resulting in increased lactic and carbonic acid at the same Pθ2, hemoglobin releases
Po2 (mmHg)
Fig. 14.10.
Bohr effect. Increases in temperature, decreases in pH, or increases in blood Pco, shift the oxygen-hemoglobin dissociation curve to the right.more O2 at that site. Mechanistically, this occurs because the binding of H+ to hemoglobin causes a conformational change decreasing hemoglobin's O2-Carrying capacity. This shift of the curve to the right is termed the Bohr effect. The reverse reaction can also occur in which binding of O2 to hemoglobin causes the liberation of H+. Decreasing the acidity shifts the oxygen-hemoglobin dissociation curve to the left.
2. Temperature. An increase in temperature also shifts the oxygen-hemoglobin dissociation curve to the right. Like increased acidity, increased temperature is a by-product of increased metabolism. Increased metabolism requires additional O2, so shifting this curve to the right provides necessary O2.
3. 2,3-Bisphosphoglycerate (BPG). An increase in the production of BPG, formerly called diphosphoglycerate, also shifts the curve to the right, thus liberating more O2. Increased production of BPG is also associated with increased metabolism.
4. Pco-,- A decrease in pH acts similarly to an increase in Pqo-,- As shown next, CO2 can react with water to form carbonic acid, which then dissociates to form bicarbonate and H+. Thus, increased Pco-, is associated with decreased pH.
Hemoglobin-nitric oxide
Nitric oxide (NO), a gas, plays an important role in vasomotor tone. It is a potent vasodilator. Produced in lung and endothelial cells, NO can be carried by hemoglobin. The binding of O2 to hemoglobin causes a change in the conformation of hemoglobin allowing NO to bind to the cysteine of hemoglobin. This protects NO from being broken down by the Fe in hemoglobin. As oxyhemoglobin releases its O2, it simultaneously releases NO.
NO dilates local blood vessels, thus further aiding in supplying O2 to areas in need.Carbon dioxide transport
Carbon dioxide is a waste product of metabolism. It is transported in the blood to the lungs in three forms:
1. Dissolved CO2. Accounting for the smallest amount of transported CO2, 7-10% is carried dissolved in the plasma.
2. Carbamino compounds. Approximately 20% of CO2 is transported in the red blood cells attached to the amino acids of globin forming Carbamino- hemoglobin. Since CO2 is bound to globin, and not the heme molecule, the CO2 does not compete with O2 or NO transport:

The formation of Carbaminohemoglobin is influenced by Pco, and the degree of oxygenation of hemoglobin. In the lungs, where Pqo-, in alveolar air is low, carbon dioxide dissociates from hemoglobin and is exhaled. At the tissue level, where Pco, is high, the formation of Carbaminohemoglo- bin is favored since deoxygenated hemoglobin readily combines with CO2.
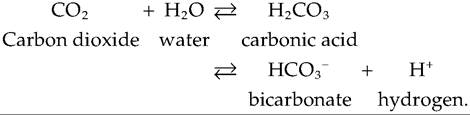
3. Bicarbonate ions. Most carbon dioxide, about 70%, is transported in the blood as bicarbonate ions (HCO3 ) (Fig. 14.11). As tissues produce CO2, it diffuses into the systemic capillaries where most of it enters the red blood cells and, in the presence of carbonic anhydrase, it combines with water to produce carbonic acid. Carbonic acid dissociates to bicarbonate and hydrogen ions. The hydrogen ions bind to hemoglobin, causing the Bohr effect and enhancing the release of O2. HCOy diffuses from the red blood cells into the plasma and is transported to the lungs. As HCOy leaves the red blood cells, chloride ions (CL) enter, a process called the chloride shift.