VIRAL INFECTIONS
DNA Viral Infections
Guinea Pig Adenovirus Infection
Outbreaks of respiratory disease attributed to guinea pig adenovirus (GPAdV-1) have been recognized in Europe, North America, and Australia, and probably occur elsewhere in the world.
The disease is characterized by low morbidity, but the mortality rate in clinically affected animals may reach 100%. In those cases described to date, animals have often been subjected to experimental manipulations that may have resulted in impairment of the immune response. GPAdV infections in colonies of guinea pigs are probably more prevalent than generally recognized. Clinical disease appears to occur primarily in young animals. Typical lesions have been observed in the airways of clinically normal young adults, emphasizing that subclinical infections do occur.Pathology
Consolidation of the cranial lobes of the lung and hilus is a characteristic finding at necropsy. Microscopic changes are those of a necrotizing bronchitis and bronchiolitis, with desquamation of lining epithelial cells and leukocytic infiltration, mononuclear cells predominating. Some airways may be obliterated by cell debris, leukocytes, and fibrinous exudate. Numerous necrotic foci may be scattered throughout the lung. The nuclei of affected epithelial cells often contain round to oval basophilic inclusion bodies 7-15 μm in diameter (Fig. 5.10). The virus has not been recovered and characterized to date, but electron microscopic examination has revealed typical adenovirus particles in affected nuclei. Using homogenates of lung prepared from a spontaneous case of the disease, typical lesions have been produced in intranasally inoculated newborn guinea pigs. The incubation period was 5-10 days. Older inoculated animals were relatively refractory to the disease.
Diagnosis
The presence of necrotizing bronchitis and bronchiolitis in young guinea pigs with characteristic intranuclear basophilic inclusion bodies is consistent with adenoviral
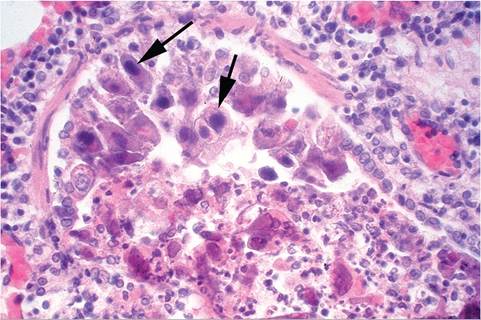
FIG.
5.10. Lung from a natural case of adenoviral bronchoalveolitis in a young guinea pig. Note the prominent intranuclear inclusion bodies (arrows) in the exfoliating bronchial epithelial cells and peribronchial leukocyte infiltration.pneumonitis. The diagnosis may be confirmed by immunocytochemistry, serology, PCR, or the demonstration of adenoviral particles in affected cells by electron microscopy. In the past, mouse adenovirus was utilized as a heterotypic antigen for serologic testing of GPAdV, but was relatively insensitive. Recently, a homotypic antigen has been engineered using the GPAdV hexon gene incorporated into replication defective adenovirus vector. This assay has proven to be more specific and sensitive. Differential diagnoses include parainfluenza virus infections, cytomegalovirus infections, and bacterial infections of the lower respiratory tract, such as Bordetella bronchiseptica.
Guinea Pig Cytomegalovirus Infection
Members of the cytomegalovirus group are species-specific viruses of the family Herpesviridae. Guinea pig cytomegalovirus (GPCMV), also known as Caviid herpesvirus 2, is quite common among conventionally housed guinea pigs, and salivary gland lesions are typically discovered as incidental findings. Guinea pigs housed under conventional conditions seroconvert to GPCMV by a few months of age.
GPCMV is transmitted by exposure to infected saliva or urine or as a transplacental infection. The latter feature has prompted interest in GPCMV as a model of congenital CMV infection. Infection may persist as an inapparent or latent infection for years. Systemic disease, with associated lesions, has been produced in weanling guinea pigs inoculated subcutaneously with GPCMV. In the experimental disease, focal lesions with intranuclear inclusion bodies were present in salivary gland, liver, spleen, lung, and kidney. Pregnant guinea pigs developed more extensive visceral lesions when inoculated with GPCMV than did nonpregnant animals. Lymphoproliferative disease, with mononucleosis-like syndrome and lymphadenopathy, has been observed in guinea pigs following experimental inoculation with GPCMV.
Infection of neonatal guinea pigs resulted in growth retardation, thymic involution due to depletion of T-lymphocytes, splenomegaly due to lymphoproliferation, and immunosuppression. However, naturally occurring GPCMV infections rarely cause detectable clinical disease in the guinea pig, a pattern similar to that observed in human CMV infections. There is 1 report of systemic CMV infection with visceral lesions in 2 young guinea pigs introduced into a conventional facility. Focal destructive lesions with large intranuclear and cytoplasmic inclusion bodies were observed in various tissues including spleen, liver, kidney, and lung. Pregnancy can precipitate acute generalized infection in sows. Both natural and experimental infections have been shown to cause abortion, stillbirth, and neonatal mortality.Pathology
Lesions are usually confined to the ductal epithelial cells of the salivary glands. Large eosinophilic inclusion
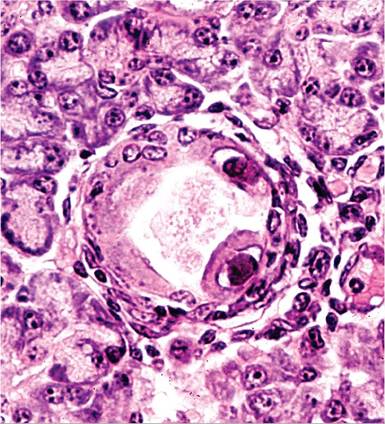
FIG. 5.11. Submandibular salivary gland from an adult guinea pig with cytomegalovirus infection. Note the karyomegaly and large intranuclear inclusion bodies, with margination of nuclear chromatin in cells lining a duct. (Source: G.D. Hsiung.)
bodies are associated with marked karyomegaly and margination of the nuclear chromatin in affected cells (Fig. 5.11). Intracytoplasmic inclusions are occasionally present in ductal epithelial cells. There may be a concurrent mononuclear cell infiltration around infected ducts. In the acute systemic form of the disease, interstitial pneumonitis with multifocal areas of necrosis in the lymph nodes, spleen, liver, kidney, lung, and other viscera are typical findings. Intranuclear and intracyto- plasmic inclusion bodies may be present in affected foci. Congenital infection can be associated with encephalitis and labyrinthitis, features that have attracted interest in GPCMV as a model of human disease.
Other Guinea Pig Herpes Viral Infections
Guinea pig “herpes-like virus” (GPHLV, Caviid herpesvirus 1) is a lymphotropic virus that was isolated from degenerating primary kidney cell cultures prepared from strain 2 guinea pigs, but to date GPHLV has not been shown to be capable of producing natural disease in guinea pigs. Guinea pig “X virus” (GPXV, Caviid herpesvirus 3) was originally isolated from the leukocytes of strain 2 guinea pigs. Based on serologic studies and DNA analyses, GPXV is different from either GPHLV or GPCMV. Following experimental inoculation of GPXV into Hartley guinea pigs, focal hepatic necrosis and mortality was observed. GPHLV and GPXV do not appear to be important primary pathogens in the guinea pig. However, they represent a possible complicating factor, should either occur as an inapparent infection in guinea pigs under experiment in the laboratory.
Equine Herpesvirus 11nfection
Neurologic signs, abortions, and stillbirths were observed in guinea pigs housed in a European zoo in association with an Equine herpesvirus 1 outbreak. The outbreak involved a variety of other species, some of which were cohoused in the same building as the guinea pigs. Nonsuppurative meningoencephalitis, neuronal and glial necrosis, gliosis, and intranuclear inclusions were observed.
Poxvirus Infection
There is a single report of 8-month-old guinea pigs within a colony that developed markedly swollen thighs due to fibrovascular proliferation. The tissue was grown in culture, and poxvirus-like structures were observed by electron microscopy. Anecdotal information among guinea pig fanciers in the United Kingdom has suggested that a poxvirus may be associated with cheilitis, but this has not been documented in the scientific literature.
RNA Viral Infections
Arenavirus Infection: Lymphocytic Choriomeningitis Virus Infection
Lymphocytic choriomeningitis virus (LCMV) infection is relatively rare in guinea pigs, but it does represent an infection that can complicate research projects and is of public health significance.
Lesions observed in guinea pigs with LCMV infection have included lymphocytic infiltrates in the meninges, choroid plexi, ependyma, liver, adrenals, and lungs. There is a wide host range, including wild mice. Exposure may occur by inhalation or ingestion, and apparently in guinea pigs through the intact skin. Confirmation requires the demonstration of viral antigen in affected tissues, serology, and/or PCR. LCMV infection has been shown to prolong the life of guinea pigs prone to L2C leukemia, which emphasizes the potential for the virus to be an important complicating factor in certain types of research. Several species, including humans, are susceptible to LCMV infection.Coronavirus-Like Infection
A syndrome characterized by wasting, anorexia, and diarrhea has been observed in young guinea pigs following their arrival at a research facility. The disease was characterized by low morbidity and mortality. Affected animals had an acute to subacute necrotizing enteritis involving primarily the distal ileum, with copious amounts of mucoid material present throughout the gastrointestinal tract. On microscopic examination, lesions were particularly prominent in the terminal small intestine. There was blunting and fusion of affected villi, with necrosis and sloughing of enterocytes from the tips of villi and formation of epithelia syncytia in the intestinal mucosa. Viral particles consistent with the morphology of a coronavirus were demonstrated in fecal samples examined by electron microscopy. In another study, clinically normal guinea pigs of different ages were observed to shed coronavirus-like particles in the feces for long periods of time. The importance of suspected coronavirus infections in this species is currently unknown. However, until additional information is available, it should be considered in the differential diagnoses in cases of enteritis and/or wasting in young guinea pigs.
Influenza Virus Infection
Influenza viruses belong to the family Orthomyxovir- idae, and are divided into 3 antigenic types: A, B, and C.
Guinea pigs are experimentally susceptible to unadapted human influenza A and B viruses, and readily transmit infection among one another under both direct and indirect contact conditions. Experimentally induced lesions tend to be mild, consisting of rhinitis, tracheitis, bronchitis, and alveolitis. A serosurvey performed among farmed guinea pigs in Ecuador revealed a high prevalence of seroconversion to both influenza A and B viruses. Although naturally occurring clinical disease has not been reported, guinea pigs appear to be susceptible to natural infection with influenza viruses.Parainfluenza Virus Infections
The family Paramyxoviridae contains a number of viruses that infect guinea pigs, including multiple members of the subfamily Paramyxovirinae: murine parainfluenza virus-1 (Sendai virus), parainfluenza virus-2 (Simian virus-5, SV-5), human parainfluenza virus-3 (PIV-3), guinea pig parainfluenza virus-3 (GpPIV-3), Cav- iid parainfluenza virus-3 (CavPIV-3), and 1 member of the subfamily Pneumovirinae: Pneumonia Virus of Mice (PVM). Much of the documentation on these infections in guinea pigs is based upon natural seroconversion, but antigens are notably cross-reactive among these viruses. None of these agents are known to cause natural clinical disease in guinea pigs. Sendai virus, SV-5, and PIV-3 have all been isolated from naturally infected guinea pigs.
Based on serologic surveys, PIV-3 antibodies are relatively common in colonies of guinea pigs. Newborn animals born to PIV-3-positive sows acquire protection via maternal immunity during the first 2 weeks of life, then usually become transiently infected (based upon seroconversion) anywhere from 2 to 8 weeks of age. Two isolates of PIV-3 (GpPIV-3 and CavPIV-3) have been isolated from subclinically infected guinea pigs, genetically sequenced, and are closely related to human and bovine PIV-3s, as well as to each other. It is unclear if these viruses are of human origin. Experimental inoculation of guinea pigs with CavPIV-3 resulted in seroconversion, but there is no clinical or histological evidence of the disease. In guinea pigs inoculated experimentally with human PIV-3, animals developed a transient interstitial pneumonia and alveolitis, with pulmonary congestion and hemorrhage. Residual lesions persisted for up to 50 days. In addition to interstitial alveolitis, documented changes have included increased histamine release from basophils, and decreased phagocytic activity of heterophils. Guinea pigs infected with human PIV-3 have been used to study mechanisms of virus-induced asthma and airway hyperactivity.
Picornavirus Infection
Antibodies to murine encephalomyelitis virus (MEV) are occasionally observed when guinea pigs are tested for this virus. Weight loss and paresis have been noted in guinea pigs that were seropositive to MEV, and meningoencephalitis was observed. Clinical disease in seropositive pet guinea pigs was also attributed to MEV infection, although the animals recovered following treatment with vitamin C. The significance of seroconversion to MEV in this species has not been resolved.
Rabies Virus Infection
A single case of raccoon-variant rabies virus infection has been documented in a pet guinea pig, requiring postexposure treatment of humans.
Other Viral Infections
Serology studies indicate that guinea pigs will sero- convert to reovirus 3.
Endogenized Viruses
Guinea pigs have an endogenous retrovirus known as guinea pig retrovirus (GPRV). GPRV has also been called guinea pig C-type virus, but it is morphologically similar to murine B-type viruses. It is serologically distinct from mouse, rat, and hamster retroviruses. Expression has been observed in cells and tissues of L2C leukemic guinea pigs and cultured cells and cell lines from other guinea pig strains. Although it is claimed to be oncogenic, its association with leukemia is coincidental and efforts to induce leukemia with GPRV have failed. Like other mammals, the genome of hystrico- morph species contains a conserved endogenous retroviral syncytin-like env-Cavl, which is expressed at the level of the placental invasive trophoblast junctional zone. In addition to endogenous retrovirus integration, the guinea pig genome contains a number of incomplete RNA and DNA viral sequence integrations, including bornavirus-, parvovirus-, and filovirus- related sequences. None of these elements have clinical significance.