VIRUSES MAINTAINED BY HOSTS WITH CLINICALLY ACTIVE INFECTIONS
RABIES VIRUSES
causative agents Rabies is one of the most lethal diseases affecting mammals and is one of the first recognized diseases (Steele and Fernandez 1991); descriptions ofrabies-like symptoms are reported from ancient Greece prior to 2300 B.C.
(Rupprecht et al. 2001, Bowen 2011b). This severe disease is caused by viruses in the genus Lyssavirus, Family Rhabdoviridae (App. 1: Table 8). Newly recognized “rabies-related” lyssaviruses have been reported in recent years, including several lyssaviruses from bats in Australia, Europe, and Africa (Acha and Szyfres 2003b, Fooks et al. 2003, Bowen 2011b). However, here we address only the classical rabies virus (Lyssavirus serotype/genotype 1) (Blanton et al. 2012).The genome of rabies viruses is a single strand ofnegative-sense RNA. Each virion is bullet shaped, with a helically coiled nucleocapsid surrounded by a double-layered lipid envelope. Replication takes place in the cytoplasm of the host cell, and newly transcribed viral genomic RNA is assembled into a nucleocapsid by helical wrapping of the protein coat (Acha and Szyfres 2003b, Bowen 2011b). Once assembled, the rabies nucleocapsids bind to the inner plasma membrane and bud through the membrane, creating the double-layered envelope. It is important to recognize that there are host-associated strains that differ in the structure of two of the five viral proteins associated with each virion: the RNA nucleoprotein and the glycoprotein that forms the envelope spikes (Rupprecht et al. 2001, Acha and Szyfres 2003b, Bowen 2011b).
host range and geographic distribution All mammals appear to be susceptible to rabies, but carnivores and bats typically serve as the chief hosts and as sources of the virus for other mammals including livestock, pets, and humans (Baer 1991, Rupprecht et al. 2001). There is one unusual case of a great horned owl (Bubo virginianus) that was infected under laboratory conditions; it shed the virus but did not develop signs of disease (Jorgenson and Gough 1976).
Rabies can be found on all continents and many islands, with the notable exceptions of New Zealand, Australia, Hawaii, and the Caribbean, where it apparently has never occurred, as well as Japan and the United Kingdom, where it has been eliminated (Rupprecht et al. 2001).
reservoirs and transmission Transmission of rabies virus is direct, typically through saliva, involving the bite or scratch of an infectious animal; in these cases transmission is believed most efficient during manifestation of the behavioral neurologic signs of disease. However, rabies viruses sometimes can be transmitted by ingestion (Correa-Giron et al. 1970, Bell and Moore 1971) or by aerosols in caves harboring large numbers of bats (Constantine 1967).
In rabies epizootiology, the term reservoir applies specifically to host species that maintain the disease in nature; thus, while virtually all mammals are susceptible, only a few species serve as reservoirs (Rupprecht et al. 2001). Rabies in domestic dogs (Canis famil- iaris), a common reservoir in many regions, has achieved virtual worldwide distribution, usually with human contributions to its spread (Smith and Seidel 1993, Knobel et al. 2005).
In the United States, wildlife accounts for over 90% of reported rabid animals, with the primary reservoir species being raccoons (Pro- cyon lotor), bats (Order Chiroptera), skunks (Mephitis mephitis, Spilogale spp.), foxes (Vulpes vulpes, Urocyon Cinereoargenteus, Alopex lagopus), and coyotes (Canis latrans) (Blanton et al. 2012). While all rabies strains are probably infectious for all mammals, each viral strain is associated with, and apparently adapted to, ecological maintenance within specific, regionally important host species (Rupprecht et al. 2001, Centers for Disease Control 2012a). Thus, skunk rabies occurs in California and the Great Plains regions, raccoon rabies in the eastern United States, Arctic fox rabies in Alaska, and coyote (Canis latrans) and fox rabies in Texas (Blanton et al.
2012). In the United States, the relative importance of different wildlife species in the maintenance of rabies has varied over time.Rabies virus strains (as well as other lyssaviruses) have foci and reservoirs among wildlife in many other parts of the world. These include fox (V. vulpes), raccoon dog (Nyctereutes procyo- noides), and bat rabies in Europe (Holmala and Kauhala 2006); mongoose (Family Herpestidae) rabies in Puerto Rico; and mongoose and wild canid rabies in Africa (Rupprecht et al. 2001).
A number of rabies strains are endemic among many bat species throughout the Old and New Worlds. Transmission from common vampire bats (Desmodus rotundus) is common in Latin America (McColl et al. 2000), and they are the most serious source for rabies in livestock (Meltzer and Rupprecht 1998); they also are a significant risk to humans in Latin America (Schneider et al. 1996).
Rabies virus has been isolated from 30 of 39 bat species evaluated in North America north of Mexico (Constantine 1979). Among 721 bats of seven species sampled in the United States, six species were found to include individuals with neutralizing antibodies, with prevalences ranging from 7 to 32%; thus some species of bats appear to be commonly exposed to rabies virus, but infections may fail to progress to clinical disease (Bowen et al. 2013). In the United States, 26 of 29 indigenously acquired human rabies cases between 1980 and 2000 were bat- associated strains, even though bat bites were documented in only two of these 26 cases; bat rabies variants identified were associated primarily with silver-haired bats (Lasionycteris noctivagans) and eastern pipestrels (Pipistrellus subflavus) (Jackson and Fenton 2001).
In contrast, bats have been associated with very few human cases of rabies in Europe. It is misleading to combine all species of bats into a single risk category because only a few bat species have been associated with most cases of human rabies (Olnhausen and Gannon 2004, Rupprecht et al.
2006).CLINICAL EFFECTS AND IDENTIFICATION Rabies is an acute, almost invariably fatal, inflammation of the lining of the spinal cord and brain (encephalomyelitis) of mammals. Virus in the saliva of the infectious animal is introduced via a bite wound or across a mucous membrane and crosses into the neural axons of both motor and sensory nerve cells. The virus particles are moved passively toward the central nervous system (CNS) (Rupprecht et al. 2001).
Once in the CNS, rabies virus particles move to specific neural sites, including deep cerebellar nuclei, cerebral cortex, brainstem, and the limbic system, as well as the thalamus and pyramidal neurons of the hippocampus (Rupprecht et al. 2001). Viral assembly reaches a maximum in the CNS, and infectious virions escape the neurons by budding through the plasma membranes of infected cells. Rabies viruses replicate in cell bodies in the brain; mature virions accumulate in peripheral tissues among cells innervated by the axons and dendrites of the infected neural cells such as the salivary glands, cornea, skin, and oral mucosa (Rupprecht et al. 2001). The viral predilection for the limbic system, cerebrum, and cerebellum explains some of the classic signs of rabies, including agitation, aggression, incoordination, depression, and death.
Rabies infections can be broadly classified into phases. First is as an incubation period, a brief prodromal phase, and then a clinical phase characterized as either excitatory (furious) or paralytic (dumb) rabies. During the incubation period, virus moves into the CNS and begins to accumulate in saliva, but there are no outward signs of disease. The length of the incubation period is variable and may rarely take more than a year before signs become apparent; however, incubation periods of 10 to 90 days are most common. The prodromal phase lasts only 1 to 2 days and is associated with subtle changes in behavior; wary animals may lose their fear of humans, relatively docile animals may become aggressive, and so on (Rupprecht et al.
2001).In the furious phase of rabies, animals become restless, agitated, nervous, aggressive, and show exaggerated responses to sensory stimuli (hyperesthesia). Such animals may selfmutilate by biting or scratching themselves, or they may bite other animals unpredictably. The reduced ability to swallow during the furious phase results in animals with copious saliva, explaining the “foaming at the mouth” syndrome often associated with rabid animals. In contrast, the paralytic or dumb phase is marked by depression, convulsive seizures, coma, and respiratory arrest immediately prior to death. Cases of clinical rabies progress quickly, typically causing death within 2 to 14 days after onset of signs (Rupprecht et al. 2001).
However, this progression is not always as linear as described. The prodromal phase often is inapparent or overlooked; also, different species spend proportionately different lengths of time in the excitatory and paralytic stages of clinical rabies. Moreover, the severity and duration of the behavioral signs of clinical rabies vary with host species, strain of the virus, route of infection, site of infection, and differential accumulation of virus in different areas of the CNS (Rupprecht et al. 2001). While most infected mammals progress through disease to death, three laboratory-challenged vampire bats remained healthy for more than 2 years following inoculation and shedding of virus in their saliva (Aguilar-Setien et al. 2005).
While the classical signs of disease can be profound, there is no obvious neural pathology that can be observed grossly during a necropsy. Rather, diagnosis often relies on the finding of characteristically dense, eosinophilic inclusions in cell bodies within the CNS, especially in the Purkinje cells of the cerebellum and the pyramidal cells of the hippocampus (Rupprecht et al. 2001).
Cytoplasmic inclusion bodies containing replicating virus are called Negri bodies, and their presence in host neurons provides a definitive diagnosis of rabies in animals; however, occasional false positive and false negative inclusion bodies can occur (Rupprecht et al.
2001). The fluorescent antibody test is considered to be a rapid, sensitive, and specific diagnostic method used globally; several other tests are available (Rupprecht et al. 2001). The various viral variants associated with the major reservoirs throughout the world can be identified further with monoclonal antibody tests and polymerase chain reaction tests (Rupprecht et al. 2001)population effects A devastating epizootic of raccoon rabies occurred following translocations of raccoons (Procyon lotor) from a known area of enzootic rabies in Florida to West Virginia in 1977 (Centers for Disease Control
2000). The translocations were to bolster the raccoon population for purposes of sport hunting. The incidence of rabies cases spread outward as classic epizootic waves that were traced using genetic variation in the virus, and by waves of mortality recognized in raccoons and other carnivores. Although these strains of rabies were absent from the northeastern United States prior to the translocations, raccoon-associated rabies has since expanded across the Eastern Seaboard to the Ohio Valley and into southern Canada (Map 10.2), and more raccoon cases have been reported between 1990 and 2011 in North America than for any other wildlife species (Fig. 10.3) (Blanton et al. 2012).
In other examples, rabies likely has spread with translocated coyotes (Centers for Disease Control 1995), mongooses (Rupprecht et al.
2001), and intercontinentally with domestic dogs (Rupprecht et al. 2001). There is a growing literature on the potential risks associated with wildlife translocations (Davidson et al. 1992, Woodford and Rossiter 1993, Cunningham 1996, Leighton 2002, Mathews et al. 2006), and these costs should motivate wildlife managers to consider disease issues before any translocations of wildlife.
special problems The almost invariably fatal outcome following untreated human infections, risks to and from peridomestic pets, and broad reservoirs in wild carnivores and bats result in considerable human fear and tremendous efforts to reduce human risks (Rupprecht et al. 2001, Knobel et al. 2005, Rupprecht et al. 2006). There are an estimated 24,000 to 93,000 annual cases of human
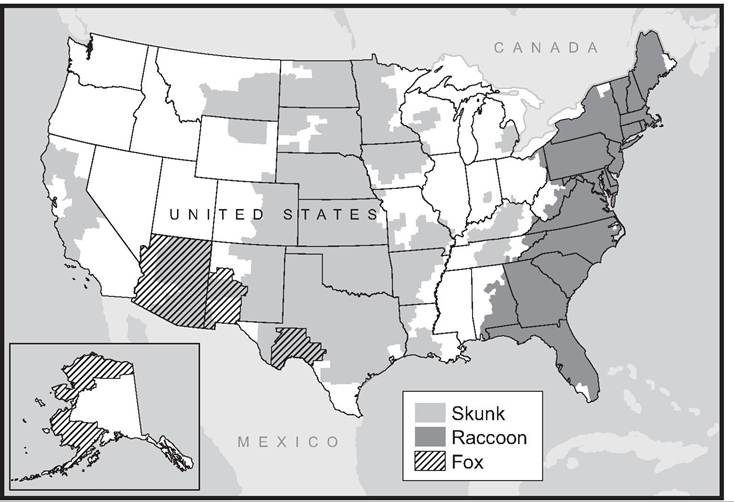
MAP 10.2. Distribution of reported rabies infections among chief wildlife hosts in the United States in 2011; note that distributions change somewhat each year and that bat-transmitted strains occur throughout much of the country (courtesy of Centers for Disease Control and Prevention, www.cdc.gov/rabies/exposure/animals/wildlife_reservoirs.html).
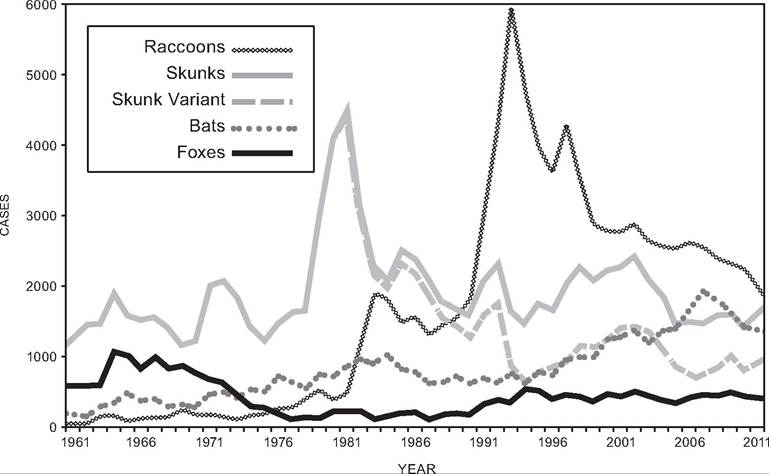
FIGURE 10.3. Annual variation in the numbers of rabies infections in the United States reported from chief wildlife host groups; multiple species of bats, foxes and skunks have been combined to simplify this graph (courtesy of Centers for Disease Control and Prevention, www.cdc.gov/rabies/resources/publications/2011-surveillance/rabid-wild-animals.html).
rabies globally, with most cases occurring in Africa and Asia, and with greater than 90% of transmission associated with bites of domestic dogs (Knobel et al. 2005). The risks of zoonotic transmission of rabies from dogs are especially great in regions where vaccination of dogs is not rigorously pursued. However, within North America, state-mandated rabies vaccination programs directed at dogs (but not at cats) have resulted in rabies being recognized in cats more commonly than in dogs or other domestic animals (Centers for Disease Control 2005, 2011).
control There is general agreement that wildlife agencies should engage in rabies control (MacInnes and LeBer 2000). General approaches for controlling wildlife rabies include elimination of the reservoir (culling), elimination of infections in the wildlife reservoir by oral vaccination, and vaccination of the potential victim species (including livestock, pets, endangered wildlife, and people) (Rupprecht et al. 2001).
For many years, culling of infected or susceptible wildlife populations was the method of choice. However, this method now is less effective than oral vaccination (Holmala and Kauhala 2006), and oral rabies vaccination with recombinant-virus oral rabies vaccine in edible baits has become the primary means of rabies control for wild carnivores (Slate et al. 2005). Evidence supports the success of oral baits in reducing wildlife epizootics (Sidwa et al. 2005), although there is some question as to whether this impact truly is cost-effective (Meltzer and Rupprecht 1998).
Control of rabies transmitted by vampire bats has involved both culling of bats and vaccination of cattle and other high-risk hosts. Oral vaccines have been considered for bats, and some work has been reported (Aguilar-Setien et al. 1998, 2002; Almeida et al. 2008), but it is difficult to translate the results into an effective vaccine (Slate et al. 2005). Vaccination of cattle is an effective means of controlling losses to rabies from vampire bats (Arellano-Sota 1988b), but it is costly (Meltzer and Rupprecht 1998). It also is argued that bat control measures are more effective in reducing rabies in cattle than vaccination (Massad et al. 2001). Vampire bat control has included use of anticoagulants such as diphenadione and warfarin (Arellano-Sota 1988a), as well as the phytoestrogen coumestrol (Serrano et al. 2007).
Reduction of human risks typically involves some combination of public education, removal of stray dogs and cats, immunization of pets and people at risk, routine diagnostic evaluation of wildlife that bite people, post-exposure treatment with immunoglobulin of unvaccinated people bitten by suspected rabid animals, and sometimes immunization of wild populations in proximity to humans to reduce epizootic risk. If a person is bitten by a domestic dog or cat appearing healthy at the time of biting, the pet can be confined by the owner for 10 days to identify the risk of exposure to rabies virus and to allow post exposure prophylaxis prior to the onset of clinical rabies. (Centers for Disease Control 2011).
CANINE DISTEMPER VIRUS
causative agent The genus Morbillivirus is in the family Paramyxoviridae. These are small, enveloped, negative-sense RNA viruses (App. 1: Table 8). Morbilliviruses include canine distemper virus (CDV) (Williams 2001), phocine distemper virus (PDV) (Kennedy 2001), measles virus (MV) (Munson 2001b), rinderpest virus (RPV) (Rossiter 2001b), peste-des-petits-rumi- nants (ovine rinderpest) virus (PPRV) (Rossiter 2001a), and the human measles virus (MacLach- lan and Dubovi 2011c, Quinn et al. 2011). Phocine distemper virus and CDV are quite closely related to each other (Blixenkrone-Muller 1993, McIlhatton et al. 1997), and Arctic seals may have been infected by CDV several hundreds or thousands of years ago by terrestrial carnivores, after which the virus evolved into PDV (Barrett 1999). Although PDV has caused several serious epizootics and infected a variety of marine mammals (Kennedy 2001, Harkonen et al. 2006), the following discussion focuses on CDV.
host range and geographic distribution Canine distemper virus occurs worldwide and has the potential of causing large-scale mortality events in a wide variety of species (Williams 2001). Severe disease develops in susceptible canids; CDV and rabies are two of the most important viral diseases afflicting this family of carnivores. However, CDV has been reported in all major families of terrestrial mammalian carnivores; in addition to Canidae, it has been reported from Felidae (wild cats), Hyaenidae (hyaenas), Mustelidae (weasel-like carnivores), Procyonidae (raccoon-like carnivores), Ailuridae (red pandas), Ursidae (bears), and Viverridae (civet-like carnivores) (Deem et al. 2000). Mortality varies among species and families; among canids 25-75% of infected animals often survive. In some families, including the mustelids, procyonids, and red pandas, mortality often is higher than typically seen in the Canidae (Deem et al. 2000, Williams 2001); nearly 100% of exposed black-footed ferrets (Mustela nigripes) and domestic ferrets (Mustela putorius furo) die (Williams 2001). Other taxa, including skunks and bears, have developed clinical distemper in captivity but seem more resistant than other carnivores (Williams 2001). Some felids, including African lions, also are susceptible. (Munson 2001a). Additionally, mortality events associated with CDV, as well as PDV, have been reported from marine mammals (Mamaev et al. 1995, Barrett 1999).
RESERVOIRS AND TRANSMISSION The severity of diseases caused by this group of viruses is directly related to the quantity of virus shed, and hence to production of acute disease. Although some recovered carriers may shed virus for several weeks to months, neither latency nor apparently healthy chronic carriers are important to the ecological maintenance of these viruses.
Transmission of CDV primarily occurs by aerosol or contact with oral, respiratory, or ocular fluids or other exudates containing the virus; close association between affected and susceptible animals is necessary because CDV is fragile in the environment (Williams 2001).
clinical effects and diagnosis Canine distemper virus enters the epithelium of the upper respiratory tract and multiplies in macrophages in infected lymph nodes before disseminating to epithelial cells throughout the body. Infected macrophages and lymphocytes often merge with other infected or noninfected cells to form nonfunctional, multinucleate, giant cells referred to as syncytia. These giant cells become factories for virus replication, and they effectively reduce the numbers of functional leukocytes. Destruction of B - and T-lymphocytes in lymph nodes, and of precursors in bone marrow, causes profound leukopenia and immunosuppression. Of course, immunosuppression makes animals more susceptible to reactivation of latent infections and to colonization by other infectious agents, including viruses (e.g., parvoviruses, herpesviruses, etc.), bacteria (e.g., Salmonella spp., secondary bacterial infections, etc.), and protozoa (e.g., Toxoplasma gondii, Pneumocystis carnii, Sarcosystis, coccidiosis, etc.) (Williams 2001). Host immunity also determines the severity of clinical distemper; hosts that develop strong responses typically survive, but those that do not may progress to severe disease and death (Williams 2001). Viral destruction of gut epithelial cells may cause hemorrhagic enteritis with explosive diarrhea and resulting dehydration. Other common signs of disease include fever, anorexia, bronchopneumonia, ocular and nasal discharge, rash and hyperkeratosis (thickening of the skin) of the epithelium of the foot pads, muzzle, eyelids, and anus, encephalitis with neuronal necrosis, depression, seizures, and death (Deem et al. 2000, Williams 2001). Distemper is often more severe, with higher levels of mortality, in juveniles than in adults. Carnivores that survive distemper may suffer lingering effects from damage to organs long after the virus is eliminated from the host, and such conditions may limit survival and fitness in the wild. Those that recover from clinical illness may continue to shed virus for up to two to three months (Deem et al. 2000, Williams 2001).
population effects Epidemics of canine distemper are maintained by a high density of susceptible hosts. However, since the host range is fairly broad, distemper epizootics can be maintained by communities of sympatric carnivores even though a given susceptible species may be rare.
Although traditionally not considered a disease of felids, clinical distemper was recognized as a problem of large cats (Panthera leo, P. tigris, P. onca, P. pardus, and P. uncia) housed in zoos in both the United States and Canada (Munson 2001a). Additionally, two wild bobcats, Lynx rufus, were diagnosed as suffering CDV-associated encephalitis. In 1994, an epizootic of CDV swept through the carnivore populations of the Serengeti-Masai Mara ecosystem in East Africa. Previous epizootics, through 1991, had killed dogs, jackals, foxes, and wild dogs without killing felids. Unexpectedly, the epizootic in 1994 killed approximately 1,000 African lions, or nearly one-third of the lion population, as well as untold leopards, hyenas, and canids (Roelke-Parker et al. 1996, Munson 2001a, Guiserix et al. 2007).
The viruses isolated from felids all appear to be a variant of CDV; the ability to cause disease in felids results from approximately 10-15% difference in the amino acid coding of surface proteins that regulate infectivity (Munson 2001a). Disease in felids is similar to that in other carnivores, but neurologic signs were common in the epizootic in 1994. Following the epizootic, approximately 85% of the two-thirds of the population that survived were shown to have been exposed, illustrating that many lions did survive exposure. However, most of those that developed clinical distemper probably did not recover. The cause of death of affected Serengeti lions ranged from virus-associated pathology such as central nervous system disease, respiratory disease, or enteritis, to a variety of secondary infections and predation by other carnivores (Roelke-Parker et al. 1996, Munson 2001a), illustrating the difficulty of assigning a single cause of death when an animal is suffering from a debilitating, immunosuppressive disease.
special problems Black-footed ferrets (Mustela nigripes) provide an excellent case study for a special problem from CDV. Blackfooted ferrets utilize prairie dog (Cynomys Iudovicianus and Cynomys leucurus) burrows for cover and specialize on prairie dogs for prey. The geographic distribution of this species once included much the prairie grasslands of North America, and populations were estimated to total more than 500,000 in 1920 (Clark 1987). However, conversion of prairies to agriculture, combined with the effects of large-scale campaigns to eradicate prairie dogs, inadvertently caused the near demise of this specialist predator (Thorne and Williams 1988).
Black-footed ferrets were considered to be near extinction when a small population was discovered in South Dakota in 1964. This population dwindled, and nine ferrets were removed to attempt captive breeding. Unfortunately, four of six of these animals died as a result of vaccination against canine distemper virus; the modified live vaccine was considered safe for use in domestic ferrets but reverted to a virulent form in black-footed ferrets. The remaining captive ferrets failed to reproduce successfully and, with no known surviving ferrets in the wild, the species was considered extinct by mid-1970s (Clark 1987, Thorne and Williams 1988).
Amazingly, another surviving population of black-footed ferrets was found near Meteetse, Wyoming, in 1981. During the following three years, the population was carefully monitored, and considerable precautions were taken to avoid disease outbreaks (dogs were not allowed in the colony, researchers handling ferrets wore face masks and disinfected their hands after handling each animal, and wild ferrets were vaccinated with an inactivated CDV vaccine) (Thorne and Williams 1988). Nonetheless, epizootics of plague in the prairie dogs and canine distemper virus in the local carnivore community were discovered in 1985. These epizootics threatened to drive this last wild population of ferrets into extinction (Thorne and Williams 1988). Six ferrets were captured in the summer of 1985 to initiate a captive breeding program. At least one of these ferrets had been exposed to CDV prior to capture and they all, unfortunately, died of the disease. Six additional ferrets were brought into captivity in 1985, another żż in 1986, and the last known wild ferret was brought into captivity in 1987; these 18 captive ferrets were the last remaining individuals of the species.
control Many inactivated and modified live virus canine distemper vaccines have been developed; however, few have been tested for safety and efficacy in nondomestic animals (Williams 2001). Modified live virus vaccines in species that can be safely vaccinated induce long-lived immunity. Also, animals surviving natural infections probably have lifelong immunity to CDV (Williams 2001).