LATENT INFECTIONS (APPARENTLY HEALTHY CARRIERS)
Transmission of some viruses through populations or communities of hosts is facilitated more by persistence in carrier animals than by the production of overt disease. Of course, the propensity to cause clinical disease is not an “either-or” property of a virus; all of the examples discussed cause clinical disease in some individuals; virus populations typically exist as a variety of genotypic “types” that vary somewhat in virulence; and, as scientists, we should expect variation that occurs along continua rather than easy, dichotomous relationships.
That said, it is instructive to consider two broad categories of viruses that produce clinical disease: one in which transmission depends upon clinical disease and the other in which subclinical carrier animals are of importance to the maintenance cycles. There are many examples of viruses with reservoirs in carrier animals that often develop subclinical infections. Avian influenza viruses and hantaviruses will be used as examples in this chapter. Other viruses that won't be discussed, but which fit this group, include the retroviruses, herpesviruses, and papilloma (or warts) viruses, among many others.AVIAN INFLUENZA VIRUSES
causative agents Viruses causing avian influenza belong to the Orthomyxoviridae (App. 1: Table 8), and those infecting wildlife and domestic animals cause Type A Influenza. Type A viruses include those that cause disease in humans, horses, pigs, canids, felids, some mus- telids, and a wide variety of birds. There are two primary proteins that have traditionally been used to classify influenza viruses: hemagglutinin (H) and neuraminidase (N). Hemagglutinin is found as 16 different types and there are nine types of neuraminidase, yielding a potential for 144 different potential combinations of these two surface proteins. However, many of these combinations have been found only in aquatic or marine birds, and most tend to be associated with asymptomatic to mild disease; a fairly small number of influenza viruses have been associated with severe disease outbreaks and deaths (Stallknecht et al.
2007, MacLachlan and Dubovi 2011b). Readers may recall the highly pathogenic avian influenza virus reported in 2012, which has a designation H5N1, as well as the 2009-2010 swine flu epidemic in North America caused by virus designated as H1N1. Perhaps more important than the pheno - typic expression of these two proteins are the sequences of eight genes that can be used to more fully characterize influenza viruses. Virulence of any given combination (such as avian influenza, AI, H5N1) can vary from strains of low pathogenicity (LPAI) to highly pathogenic strains (HPAI) that cause severe disease and death (Stallknecht et al. 2007, MacLachlan and Dubovi 2011b, Quinn et al. 2011). Virulence, including designation as HPAI, is determined based on the tendency to kill chickens (MacLach- lan and Dubovi 2011b, Quinn et al. 2011), which may or may not be associated with virulence in wild birds or mammals (including humans).Influenza viruses evolve quickly because of common rearrangement among the six to eight strands (Influenza A viruses have eight strands) of negative-sense RNA that comprise the genome. Evolutionary changes in these viruses are accomplished by two separate, but synergistic, processes (Chen et al. 2004, MacLachlan and Dubovi 2011b). Genetic drift is similar to random changes in any genome that occurs with point mutations within the various genes. Genetic shift occurs with rearrangement of genes along the strands of genomic RNA, causing dramatically rapid change in the circulating viral populations, and this is most likely with mixed infections of different strains. Thus, unlike most viruses and many other pathogens, previous infections confer long-term immunity only to the infecting strain, and, because rapid evolution leads to changes in the proteins of the virion, this does not result in long-term immunity to influenza viruses in general (Chen et al. 2004, MacLachlan and Dubovi 2011b).
The virion shape of influenza viruses is pleomorphic, ranging from filamentous to spherical, and each virion is surrounded by a lipid envelope derived from the host cell membrane.
The H and N proteins occur as spikes that extend outward from the envelope and provide the main antigenic moieties characteristic of each strain. Virulence of influenza viruses depends upon post-translational cleavage of the hemagglutinin molecule. Such cleavage is more rapid and the virus is more virulent when the cleavage site is longer; this is because more basic amino acids at this site make it easier for the enzyme to cleave the hemagglutinin. Cleavage occurs more slowly when the cleavage site is composed of a single arginine and results in strains of low pathogenicity). Thus, the H and N number designations help classify influenza viruses but they are not the sole determinants of virulence (Stallknecht et al. 2007, MacLachlan and Dubovi 2011b, Quinn et al. 2011).HOST RANGE AND GEOGRAPHIC distribution Type A influenza has been isolated from a wide variety of birds and mammals, including humans, canids, felids, mustelids, horses, pigs, chickens, turkeys, and a wide variety of wild birds, but most strains of the virus tend to be fairly specific to species or families of hosts. Wild birds commonly are infected with influenza viruses that cause little or no sign of disease and that serve as the source for most viral strains, probably including all strains of AI (Stallknecht et al. 2007, MacLachlan and Dubovi 2011b, Quinn et al. 2011). Waterfowl, gulls, and shorebirds often are infected, but they typically serve as asymptomatic carriers of a wide variety of AI strains. These strains vary between years and regions because of the dynamic evolution of the viruses. Wild birds move virus along all of the major flyways and throughout much of the world; evidence of AI has been found on all continents in areas where reservoir hosts occur.
Most influenza viral isolations have come from ducks and geese (Order Anseriformes). However, other species of birds also are susceptible to infection by avian influenza viruses; over 100 species from 13 orders, including perching birds, raptors, owls, game birds, coots, loons, and seabirds, have been found infected (Stallknecht et al.
2007).RESERVOIRS AND TRANSMISSION Water birds serve as reservoirs for most influenza strains on a worldwide basis. Low-pathogenicity AI viruses typically replicate in gut cells and are shed via feces, and transmission between birds is typically via a fecal-oral route. However, some strains, including HPAI H5N1, appear to replicate in the trachea as well as in the gut, and this clearly facilitates respiratory spread (Chen et al. 2004, Sturm-Ramirez et al. 2005, Boyce et al. 2009, Henaux and Samuel 2011). In one study, virus was shown to have been shed from ducks, but shedding ranged from 4.5 days to 18.5 days, depending on the duck species and the strain of the virus, with HPAI H5N1 shed for a shorter duration than LPAI strains (Henaux and Samuel 2011). Because of their envelopes, influenza viruses do not persist for extended periods of time in the environment due to sensitivity to low pH, heat, dehydration, and so on, and therefore persistence of the viruses in dry environments is generally short. However, in wetlands, where most virus from waterfowl is shed, infectious viral particles do persist for extended periods of time (Henaux et al. 2012). Stallknecht et al. (2007) reviewed the persistence of AI in wetlands and noted that in some circumstances AI viruses can remain infectious in water for 32 days. In colder waters of the breeding grounds of many species of waterfowl, AI viruses may persist for longer periods, providing an environmental source to both intra- and interspecific transmission. Although AI viruses persist for shorter periods of time in warmer water, Henaux et al. (2012) were able to use viral isolation and PCR to identify AI from fecal samples and water samples from summer wetlands in California. Although prevalence in waterfowl was low, and AI viruses were not isolated from water, the authors clearly showed that transmission on summer wetlands is part of the complex epidemiology of avian influenzas.
Avian influenza strains historically have remained in wild avian communities, with occasional spill-over into domestic ducks and geese, as well as domestic poultry.
The most recent pandemic of highly pathogenic avian influenza presumably began in Asia, where small numbers of wild birds were found dead during 2002 through 2004. In 2005, over 6,000 wild water birds, including bar-headed geese (Anser indicus), two species of ducks, brown-headed gulls (Chroico- cephalus brunnicephalus) and great cormorants (Phalacrocorax carbo), died on Qinghai Lake in China, and an epidemic wave spread quickly as far west as Great Britain by March of the following year (Stallknecht et al. 2007).Transmission of avian influenza viruses to chickens has been a concern of the poultry industry because epidemics of highly pathogenic “fowl plague” have caused millions of deaths (due to disease and also culling). It is also a public health concern because human infections have coincided with outbreaks in domestic poultry, and the genetics of strains isolated from humans have been identical to those in chickens (Wan et al. 2005). Transmission among avian species is facilitated by similar sialic acid receptors on the cell membranes of their intestinal cells. Humans typically are not infected directly by avian influenza viruses because our gut cell receptors are too different from those of avian cells. Pigs, on the other hand, have receptors that are similar enough to both birds and humans that they commonly serve as the bridge between birds and humans.
Rural farms in many areas of Asia provide overlap of domestic ducks that interact with wild birds on ponds, chickens in people's yards, and swine; hence, Asia often provides a source of avian influenza viruses infective to humans. Once swine become infected, genetic drift or shift that effectively allows virus to attach to swine cells provides selective advantage (Ma et al. 2009). Humans become infected from pigs once strains adapt to bind to the humanlike sialic acid receptors of pigs. Once humans are infected, further adaptations have occasionally allowed human-to-human transmission (Stallknecht et al.
2007, MacLachlan and Dubovi 2011b).The recent epizootic of highly pathogenic H5N1 differed from classic avian influenza in several ways. One important difference is that the virus evolved infectivity to humans without requiring passage through swine. Passage of the virus through swine populations often results in strains that cause adequate levels of viremia to facilitate human-to-human transmission. Death may still occur with infections causing low levels of viremia, but high levels of viremia are required of reservoir hosts capable of supporting maintenance of the virus.
The World Health Organization reported 633 human cases, with 377 confirmed deaths, of HPAI H5N1 from 15 different countries between the years spanning 2003 to July 2013 (World Health Organization 2013). The fear of a pandemic of this virus is justified because of the high case fatality rate and because the current viral type H5N1 may require only a single mutation to be transmitted from human to human (Tharakaraman et al. 2013), and because history remembers the “Spanish flu” pandemic of 1918 in which an estimated 21-40 million people died (Acha and Szyfres 2003a, Quinn et al. 2011). Luckily, the current virus strain has not evolved to yield high enough viremia or adequate shedding to allow easy transmission between humans.
cLiNicAL effects There are no clinical signs that are pathognomonic for influenza, and most birds show no pathologic signs of infection. Of course, pathology is also expected to vary by strain, host species, and individual animal characteristics associated with immune status, age, reproductive condition, nutritional status, other infections, and route of infection. Influenza viruses infect endothelial cells of the gut during early stages of infection. In domestic poultry, and in some wild birds infected with highly pathogenic strains, signs including diarrhea, respiratory distress, and central nervous system disease are common, and some of these signs have been observed in wild birds that have died of highly pathogenic H5N1. In birds that survive for more than a couple of days, disease of different organ systems is typically associated with edema and hemorrhage. Initial signs often include diarrhea, followed by respiratory disease marked by nasal discharge, coughing, sneezing, and fluids in the lungs causing rattling noises (rales) associated with interstitial pneumonia, and birds may die of pulmonary congestion, edema, and hemorrhage. Neural disease is associated with signs consistent with meningitis (inflammation of the lining of the brain and spinal cord) including tremors, torticollis (or wry neck), and opisthotonos (hyperextension of the spinal column). In severe cases in which birds survive enough days, multiple organs may show signs of hemorrhage, edema, cellular necrosis causing splenomegaly, hepatomegaly, renomegaly, pancreatic acinar necrosis, and bone marrow depletion (Stallknecht et al. 2007, MacLachlan and Dubovi 2011b, Quinn et al. 2011).
population effects Most avian influenza exists as strains of low pathogenicity, and these strains cause little or no overt disease in most wild carriers. There have been only a couple of major disease epizootics that have caused significant deaths of wild birds. The first large die-off of wild birds caused by avian influenza involved common terns (Sterna hirundo) in South Africa (Stallknecht et al. 2007), and this involved virus characterized as H5N3. The second large die-off occurred with the H5N1 outbreak of 2003-2012. Although thousands of birds might die in a major outbreak, it remains rare that these viruses cause significant population effects among wild birds.
Spillover into non-reservoir hosts, including domestic poultry, is another matter. Millions of chickens and turkeys have died from highly pathogenic avian influenza or been culled to eliminate potential reservoirs of H5N1 virus. Of course, this has caused economic hardship for many backyard breeders and cost the industry millions of dollars. In fact, since 1955, 27 epizootics of highly pathogenic avian influenza involving various strains have been responsible for the deaths of over 500 million birds, including domestic fowl that were culled (MacLachlan and Dubovi 2011b).
control Control of avian influenza viruses in wild birds seems impossible. Control in poultry and domestic ducks and geese typically involves culling as many birds as thought necessary and/or vaccination. Control of potential risks for humans is more complex. Large- scale culling efforts to rid a region of infected birds has been attempted, and domestic fowl will continue to be culled when found to be infected with HP avian influenza viruses. If the virus mutates to allow human-to-human transmission, then control of human risks will necessarily rely on vaccination programs at all scales—local individuals near cases, countries, continents, and perhaps globally.
HANTAVIRUSES
causative agent The hantaviruses belong to the family Bunyaviridae (App. 1: Table 8); this is the largest family of viruses, with at least 350 viruses recognized. Bunyaviruses are spherical RNA viruses with three singlestranded, negative-sense genomic segments. These viruses replicate in the cytoplasm and bud through both the Golgi membrane and the cell membrane. Each virion is composed of the three genomic segments, four proteins, including two external glycoproteins, and components of the two lipid layers (Bowen 2011a).
HOST RANGE AND GEOGRAPHIC distribution Hantaviruses are distributed widely throughout the world. Many of the bun- yaviruses are arthropod-borne, but the hantaviruses are transmitted from vertebrate to vertebrate without arthropod vectors. Old World Hantaan virus, Puumala virus, and Seoul virus are hantaviruses that cause hemorrhagic fevers with renal syndromes. There are more than 20 members of the genus Hantavirus in North America alone, all of them have reservoirs in rodents, and at least six cause severe disease in humans. These viruses don't cause overt disease in rodent reservoirs, but many of them cause severe disease in humans including hantavirus pulmonary syndrome (Mills et al. 2010). More than 200,000 human cases of recognized hemorrhagic fever with renal syndrome (HFRS) occur caused by Hantaan virus annually in China, Russia, and Korea, and smaller numbers of cases are reported from Japan, Finland, Sweden, France, Greece, Bulgaria, and the Balkan countries (Bowen 2011a).
An outbreak of a previously unrecognized New World hantavirus, later designated Sin Nombre virus, killed 12 of 24 human cases recognized between December 1992 and May 1993 (Mills et al. 2002, 2010). This outbreak occurred on the Colorado Plateau where the four states of Colorado, Utah, Arizona, and New Mexico meet in what is commonly referred to as the “Four Corners” region of the southwestern United States. This outbreak was different in a couple of respects from Old World Hantaan virus outbreaks. Importantly, humans developed hantavirus pulmonary syndrome (or cardiopulmonary syndrome) with severe pneumonitis rather than HFRS. Second, the initial case fatality rate was high (50%) in comparison with the 2-10% more typical for HFRS (Mills et al. 2002, 2010).
Considerable work was conducted on hantaviruses in the United States during the two decades following the 1993 outbreak in the Four Corners region. By 2010, more than 480 cases of HPS had been identified in humans from 30 states (Mills et al. 2010). Importantly, the overall case fatality rate was shown to be somewhat less than in the original outbreak, but was still very high, at 37%, when compared to most infectious diseases (Mills et al. 2002). Mills and Childs (2001) reported positive serologic results from 25 rodent species and Audubon's cottontail (Sylvilagus audubonii) to three hantaviruses, including Sin Nombre virus, indicating a wide range of exposure in small mammal communities. However, exposures say little about reservoir potential or risks of viral shedding to other reservoir hosts or humans.
RESERvoiRS and transmission Reservoirs of the Old World Hantaan viruses include field mice in the genus Apodemus, bank voles including Clethrionomysglareolus (Tesakov et al. 2010), and Old World rats in the genus Rattus (Bowen 2011a, Quinn et al. 2011, Heyman et al. 2012). Heyman et al. (2012) evaluated environmental correlates, including food availability of rodent reservoirs, carrier abundance or pathogen prevalence, rodent ecology, virus survival, local climatic conditions, and human behavior associated with human outbreaks of HFRS in Europe. Although all of these variables may influence outbreaks, the complexity of the interactions makes it difficult to accurately predict outbreaks based on any single variable (Heyman et al. 2012). These complex interactions result from a bottom-up trophic cascade in which prevalence in reservoir hosts is influenced by multiple factors associated with host abundance, which is influenced by habitat quality and food availability, which are influenced by weather and anthropogenic disturbance (Fig. 10.1).
Many rodent species in the family Muridae, including species in the genus Peromyscus, have been found to be exposed to hantaviruses; P. maniculatus (deer mouse) is generally considered the primary reservoir host of Sin Nombre virus (Childs et al. 1994; Mills et al. 2002, 2010; Bagamian et al. 2012). Clearly, P. maniculatus is the most widespread and commonly infected host (Mills and Childs 2001), and the conservative assumption is that Sin Nombre virus has the potential to occur throughout North America wherever this host
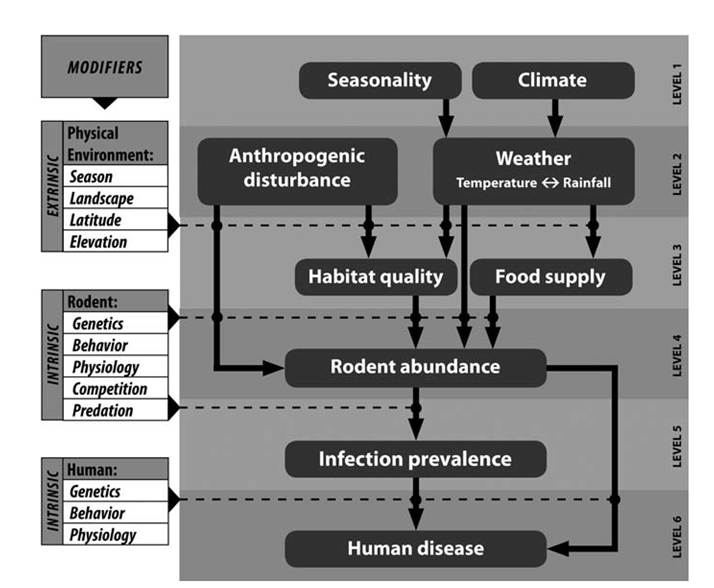
FIGURE 10.1. Simplified conceptual model of hantavirus transmission. This is a bottom-up trophic cascade in which relative risks to humans are tied to the density of reservoir hosts and the density-dependent prevalence of Sin Nombre virus among those hosts. Major influences on reservoir host abundance include habitat disturbance, habitat quality, and food supply; all of which influence reproduction, growth and survivorship. Habitat quality and food supply are influenced by habitat disturbance and local weather patterns which result from patterns of seasonality and climate. Intrinsic and extrinsic modifier variables, shown on the left, affect the cascade at the intersections of the levels within the model. Such modifiers are rarely understood and feed-back interactions should be expected among several of the modifiers as well as among some levels within the model. For example, at the level of the rodent reservoir host, abundance could conceivably affect habitat quality and food supply as well as behavior, physiology, and ultimately genetics of the rodent population (from Mills et al. 2010, copyright © Mary Ann/Liebert, Inc., Publishers, by permission).
occurs (Map 10.1). The prevalence of infections in populations of P. maniculatus has been found to vary from a few percent to 30% (Mills and Childs 2001), but the mean prevalence among most populations is approximately 10% (Mills et al. 2002). Other hosts, including P. boylii (Abbott et al. 1999), also have been shown to be exposed to hantaviruses at prevalences exceeding 10%.
Reservoirs of a variety of other recently identified New World hantaviruses vary by region and viral species. New York virus is associated with the white-footed mouse, Peromyscus leucopus. The cotton rat, Sigmodon hispidus, is a reservoir of Black Creek Canal virus. The rice rat, Oryzomys palustris, is a reservoir for Bayou virus, and there are a variety other species in the genus Hantavirus that are associated with other rodent species in Central and South America, but many of the identified viruses have yet to be associated with human disease (Zeier et al. 2005, Millws et al. 2010).
Transmission occurs as a result of exposure to aerosols or bodily fluids from infective mice, probably most importantly including infected saliva, urine, or feces (Mills et al. 2002, Zeier et al. 2005, Mills et al. 2010). Prevalence in reservoir hosts often varies with age and sex, with larger, older males typically exposed at the highest levels (Abbott et al. 1999); presumably, transmission between older, larger males includes greater sociality that results in
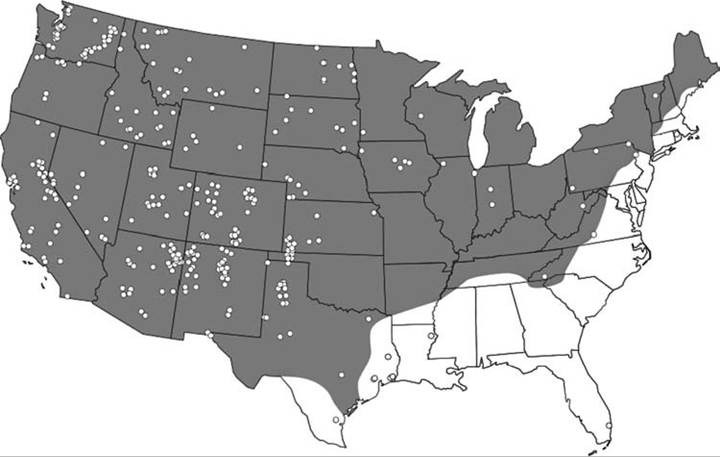
MAP 10.1. Geographic range of the deer mouse (Peromyscus maniculatus) and confirmed cases of hantavirus pulmonary syndrome (HPS) in the United States as of June 6, 2002 (from Mills et al. 2010, copyright © Mary Ann/Liebert, Inc., Publishers, by permission).
contamination of fight wounds with virusladen saliva.
Humans become infected with Sin Nombre virus most commonly when they enter and disturb enclosed spaces contaminated with urine or feces of infected mice. In most cases, humans become infected by inhalation of infectious, virus-laden aerosols; more rarely, humans become infected via exposure to broken skin, mucous membranes, rodent bites, or potentially even ingestion (Mills et al. 2002, 2010). Risks appear greatest when campers enter cabins in which deer mice have been living, but many routes of exposure are possible.
During the late summer of 2012, an outbreak of Sin Nombre virus occurred at Yosemite National Park, where at least 10 visitors were exposed and three died after staying in tent cabins (Fig. 10.2) (Centers for Disease Control 2012b). Nine of the 10 exposures occurred among people in Curry Village Signature Tent Cabins. These tent cabins have insulation between the outer tent wall and the interior wall of the cabin. Apparently, mice were nesting in the insulation and entering the cabins through small cracks associated with the doors or small gaps in the canvas. The park notified approximately 260,000 guests who had visited between 1 June and 17 September 2012. They also voluntarily closed many of the cabins indefinitely (Centers for Disease Control 2012b). Although ecological studies on the reservoir population have not been published, initial trapping successes suggested extremely high populations of P. maniculatus living in and around these cabins (Mark Novak, California Department of Public Health, pers. comm.).
Outbreaks of hantavirus have been associated clearly with peaks in population densities of reservoir hosts, and many species of small mammals experience large fluctuations in population density. Such peaks often follow years of high primary productivity, which are typically associated with years of high rainfall in arid areas of the southwestern United States (Mills et al. 2010). Although we can try to control our exposures to reservoir hosts (see below), control of wild rodent populations is difficult except at a local scale.
CLINICAL EFFECTS AND IDENTIFICATION Disease caused by the genus Hantavirus has been reported to be minimal to none in reservoir hosts, and there have been no reported

FIGURE 10.2. (A) One of at least 91 double-walled tent cabins insulated with polystyrene foam that were demolished by the Curry Village Company, Yosemite National Park, in 2012 following an outbreak of hantavirus pulmonary syndrome (HPS) that killed three people. The Curry Village Company worked with the National Park Service and California Department of Public Health, providing a good example of risk management coordinated by multiple entities working together to protect human lives. (B) A lower section of a tent cabin showing evidence of mouse entry and damage, suggesting potential exposure risks for infection by Sin Nombre virus (photos courtesy of California Department of Public Health).
population-level effects in wildlife. Reservoir hosts generally are believed to carry and transmit these viruses without developing disease (Botten et al. 2000, Zeier et al. 2005), making the hantaviruses excellent examples of pathogens that are carried and transmitted by chronic carriers; however, at least one group has shown that male P. maniculatus infected with Sin Nombre virus suffered higher mortality than did uninfected males or females (Luis et al. 2012), suggesting disease at some level.
In humans, the incubation period can range from ι to 6 weeks. Initial symptoms of hantavirus pulmonary syndrome (HPS) include fever, chills, myalgia, headache, and gastrointestinal symptoms which persist for up to a week. Following flu-like symptoms, pneumonia develops rapidly as fluids and white blood cells are shunted into the lungs, progressing very rapidly to respiratory distress, shock, and often death (Graziano and Tempest 2002). Wildlife biologists who have lived in rodent-infested cabins or trapped P. maniculatus or other reservoirs of hantaviruses, need to be aware of the signs and advocate for their own medical care when necessary. Although the risks to mammalogists and other field biologists who handle rodents in the field is apparently very low (Kelt et al. 2007), the case fatality rate remains high once signs appear, and appropriate precautions must be taken to avoid unnecessary risks.
control Mills et al. (2002) listed situations associated with higher risks of exposure ofpeople to hantaviruses. These included “increasing numbers of host rodents in human dwellings; occupying or cleaning previously vacant cabins or other dwellings actively infested with rodents; cleaning barns and other outbuildings; disturbing excreta or rodent nests around the home or workplace; residing in or visiting areas where substantial increases have occurred in numbers of host rodents or numbers of hantavirus-infected host rodents; handling mice without gloves; keeping captive wild rodents as pets or research subjects; handling equipment or machinery that has been in storage; disturbing excreta in rodent-infested areas while hiking or camping; sleeping on the ground; and hand plowing or planting in soils contaminated by deer mice.” They also reported precautions for workers frequently exposed to mice, guidelines for collecting samples, and rather extreme personal protective equipment necessary to handle mice in the field (Childs et al. 1995, Mills et al. 1995). These precautions are very thorough and justified because of the high human case fatality rate; human safety should be the first concern for all wildlife projects.
However, serologic surveys of people with occupational rodent exposure have revealed few with antibodies to hantaviruses (Fulhorst et al. 2007, Kelt et al. 2007). Fulhorst et al. (2007) reported that only four of 757 (0.53%) people whose jobs required contact with rodents were serologically positive for Sin Nombre virus. Similarly, Kelt et al. (2007) reported that only one (0.07%) of 1,412 mammalogists with exposures to large numbers of wild rodents had been hospitalized with signs consistent with HPS, and four others (0.28%) who were serologically positive for hantaviruses. This evidence of exposure suggests that most wildlife biologists who handle rodents in the field for purposes other than disease sampling are at relatively low risk of infection; of course, those working with deer mice in areas where hantaviruses are known to occur should take appropriate precautions.
Recent recommendations for mammalogists and field biologists who trap rodents for reasons other than disease surveys include a more practical list of eight guidelines for minimizing most risks of exposure to hantaviruses (Kelt et al. 2010). These guidelines include (1) avoidance of staying in cabins or field bunkhouses showing evidence of rodent occupation until the facilities are thoroughly cleaned. Cleaning should include spraying of surfaces with liquid disinfectants (10% bleach, 3% Lysol or a variety of virucides) to control dust containing virus particles. Mops should be used to remove feces rather than brooms that sweep and disperse virus into the air, and those doing the initial cleaning should wear a properly fitted HEPA filtered mask. (2) Rodents considered to be potentially infective should be handled in the open air, and traps and rodents should be held “downwind” to minimize respiratory aerosols. (3) Live traps containing live rodents or contaminated bedding should not be transported in closed vehicles without first isolating them in closed plastic bags. (4) Appropriate gloves should be worn, and mammalogists should avoid contact with feces, urine, or rodent bites.
(5) Training should be provided to all workers handling rodent reservoirs to ensure that they avoid being bitten or otherwise exposed.
(6) In the event that a field biologist is bitten, scratched, or contacts urine or feces, the area should be washed with soap and water and then sanitized with hand sanitizers or other disinfectants. (7) Field workers should be aware of the signs of infection by hantavirus, and seek medical attention when signs appear. Importantly, ill field workers should explain potential exposures to their physicians and be ready to take responsibility for their own health if their physicians assume that all flu-like symptoms are necessarily flu or when respiratory problems progress from irritating to serious respiratory problems. The importance of awareness of potential disease risks by the field biologists taking those risks cannot be overstated. (8) Finally, biologists and managers should attempt to stay up to date with information about their risks, including regular review of guidelines provided on the CDC websites.
More on the topic LATENT INFECTIONS (APPARENTLY HEALTHY CARRIERS):
- LATENT INFECTIONS (APPARENTLY HEALTHY CARRIERS)
- Carriers by Sea, Innkeepers and Stablekeepers
- Why humility matters: healthy moral functioning
- Fundamentals of healthy lifestyle
- TREATMENT OF RESIDUAL HIV-1 DISEASE: PURGING LATENT
- Jean-Leon Gerome’s The Snake Charmer (Figure 24.1) is a painting about mystery and latent danger in the exotic East.
- Other Bacterial Infections Corynebacterium spp. Infections
- Bloodstream Infections and Catheter-Related Bloodstream Infections
- Skin, Soft-Tissue, and Bone Infections Purulent Skin and Soft-Tissue Infections (Furuncles, Carbuncles, Abscesses)
- STAPHYLOCOCCUS INFECTIONS
- HERPETIC VIRAL INFECTIONS
- OPPORTUNISTIC INFECTIONS
- INTRAUTERINE INFECTIONS
- PERINATAL INFECTIONS
- NOSOCOMIAL INFECTIONS
- FUNGAL INFECTIONS
- INTRACRANIAL INFECTIONS
- BIBLIOGRAPHY FOR FUNGAL INFECTIONS
- ENTERQViRAL INFECTIONS (NON-POLIO)
- HERPESVIRUS INFECTIONS IN WILD BIRDS