Vision
Vision is a major sensation in many animals. This is reflected in the large number of receptors within the eye and the large percentage of the cortex that is devoted to vision. Vision is particularly important in birds.
Although the eyes make up only about 1% of the weight of the head in humans, they make up approximately 15% in a starling.Accessory structures of the eye
The accessory structures of the eye include the eyelids, eyelashes, eyebrows, lacrimal (tearing) apparatus (Fig. 11.7), and extrinsic muscles of the eye.
Eyelids, eyelashes, and eyebrows
The upper and lower eyelids, or palpebrae, cover the eye during sleep, protect the eye from excessive light and foreign objects, and assist with lubricating the eye (Fig. 11.8). The layers of the eyelid consist of the superficial epidermis, dermis, subcutaneous tissue, fibers of the orbicularis oculi muscle, a tarsal plate, tarsal glands, and conjunctiva. The tarsal plate is a fold of connective tissue giving support to the eyelid. Embedded in the tarsal plate is a row of sebaceous glands called the tarsal, or Meibomian, glands. They secrete
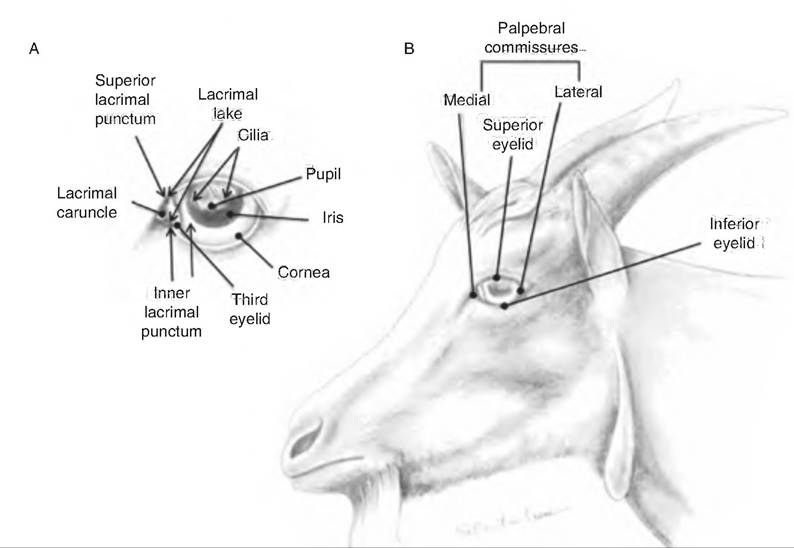
Fig. 11.7. External structures of the eye in the goat. (A) Frontal aspect of the eye and lacrimal apparatus. (B) Eyelids and eye of the goat. (Reprinted from Constantinescu, 2001. Used by permission of the publisher.)
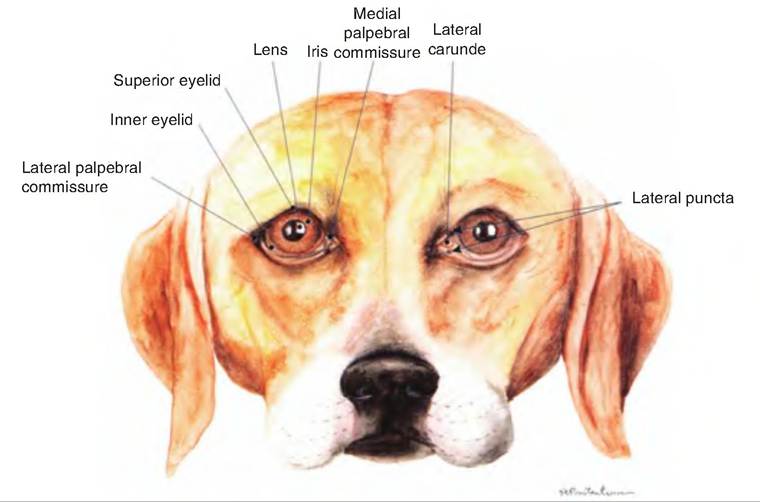
Fig. 11.8. Front view of the eye of the dog. (Reprinted from Constantinescu, 2002. Used by permission of the publisher.)
fluid that helps prevent the eyelids from adhering from one another. The conjunctiva is a thin, protective mucous membrane consisting of stratified columnar epithelium with goblet cells and areolar connective tissue.
It lines the inside of the palpebrae and the anterior surface of the eyeball, excluding the cornea.The gap between the two eyelids is the palpebral fissure. At either corner of the eyelid is the lateral and medial commissure, respectively. A small, reddish elevated area found in the medial commissure is the lacrimal caruncle. It contains both sebaceous (oil) and sudoriferous (sweat) glands.
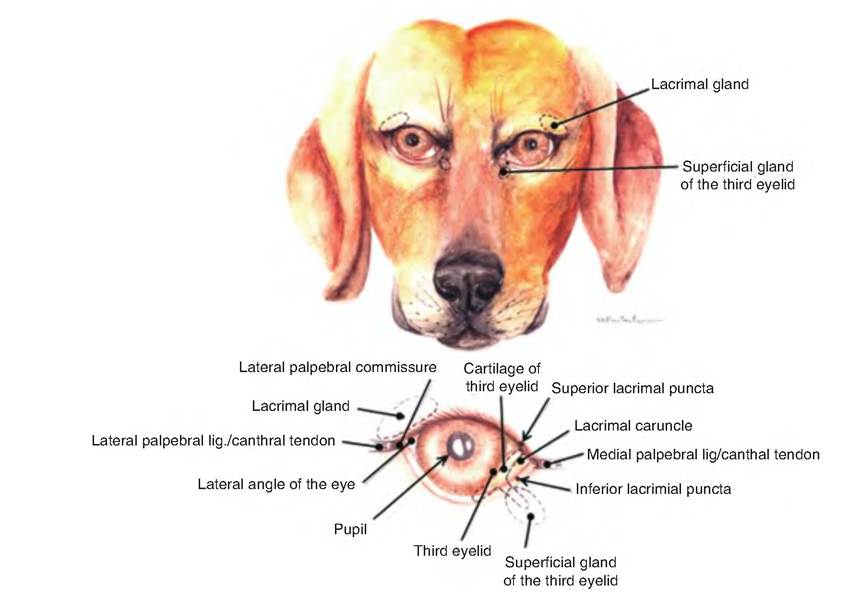
Fig. 11.9. Eye and accessory structures of the dog. (Reprinted from Constantinescu, 2002. Used by permission of the publisher.)
Domestic species have a nictitating membrane or third eyelid (Fig. 11.9). This fold of mucous membrane arises from the ventromedial border of the eye. At its base is a serous gland called the gland of the third eyelid.
Located at the margin of the eyelids are the cilia (eyelashes). Located above each eyelid are the eyebrows. Both structures help keep foreign objects, perspiration, and direct sunlight out of the eye. Located at the base of the hair follicles of the eyelashes are sebaceous ciliary glands that release a lubricating fluid. Infection of these glands is a sty.
Lacrimal apparatus
The lacrimal apparatus (Fig. 11.9) consists of a group of structures that produce and drain tears (lacrimal fluid). Lacrimal glands, located in the dorsolateral portion of the orbit, secrete lacrimal fluid through excretory lacrimal ducts that empty onto the surface of the conjunctiva of the upper eyelid. The fluid moves over the anterior surface of the eye and enters two small openings on the upper and lower palpebrae near the medial corner called the lacrimal puncta. From there, the fluid enters the lacrimal canals, two ducts leading into the lacrimal sac. The nasolacrimal duct carries fluid from the lacrimal sac into the nasal cavity just below the inferior nasal concha. Lacrimal fluid contains water, salts, mucus, antibodies, and lysozyme. This fluid protects, lubricates, and moistens the eyeball.
The moisture seen on the nose of domestic animals is largely lacrimal fluid.Extrinsic eye muscles
Movement of the eyeball is controlled by six striated muscles called extraocular muscles, in contrast to intraocular muscles located within the eyeball (Fig. 11.10). The lateral and medial rectus muscles move the eye laterally and medially, respectively. The superior rectus and inferior rectus muscles elevate and depress the eye, respectively. The inferior oblique muscle elevates and turns the eye laterally, while the superior oblique depresses and turns the eye laterally.
Anatomy of the eyeball
The receptors for vision reside within the eyeball. The eyeball consists of three layers: (1) fibrous tunic, (2) vascular tunic, and (3) retina (Fig. 11.11).
Fibrous tunic
The fibrous tunic, or external coat of the eyeball, is avascular and consists of the anterior cornea and posterior
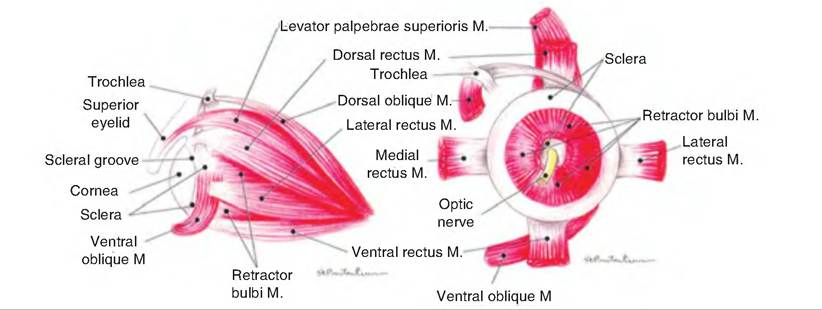
Fig. 11.10. Muscles of the eye of the dog. The left drawing shows the lateral aspect of the eye while the right drawing shows the posterior aspect of the eye. (Reprinted from Constantinescu, 2002. Used by permission of the publisher.)
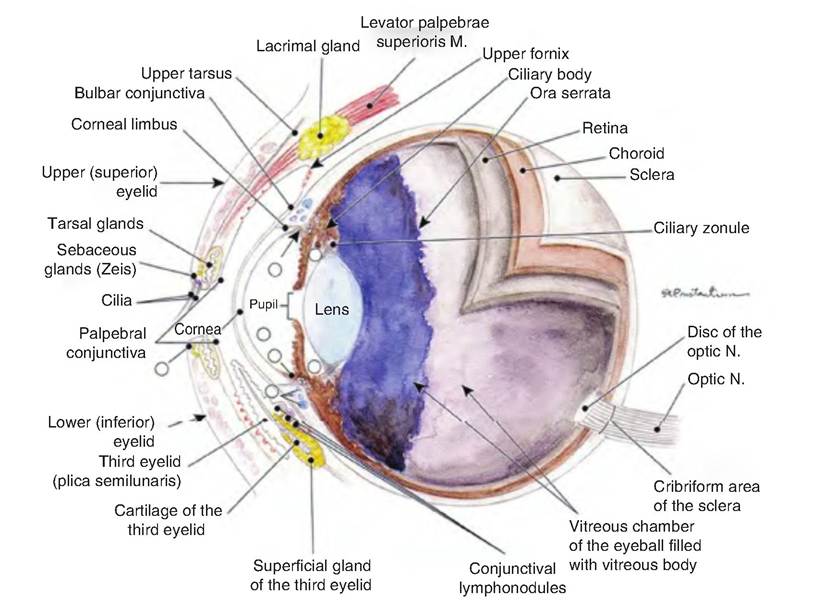
Fig. 11.11. Median section of the eye of a dog. M, muscle; N, nerve. (Reprinted from Constantinescu, 2002. Used by permission of the publisher.)
opaque sclera. The cornea is a transparent layer covering the iris, the colored portion of the front of the eye. The cornea is curved and therefore helps bend light toward the retina. The cornea consists of three layers. Its outer layer consists of nonkeratinized stratified squamous epithelium, the middle layer consists of collagen fibers and fibroblasts, and the inner layer is simple squamous epithelium. The sclera covers the entire outside surface of the eye with the exception of the cornea. It consists of dense connective tissue and provides rigidity, helping to maintain the shape of the eyeball.
The scleral venous sinus (canal of Schlemm) is located at the junction of the sclera and cornea. Aqueous humor drains into this sinus.Vascular tunic
The vascular tunic, or uvea, is the middle layer of the eyeball. It consists of three parts: choroid, ciliary body, and iris. The choroid is the highly vascularized, dark brown, posterior portion of the vascular tunic, lining the majority of the inside of the sclera. Its brown pigment is produced by melanocytes, and helps absorb light so that it is not scattered throughout the inside of the eye. The choroid is incomplete where the optic nerve exits the posterior of the eyeball.
Many species of domestic animals, including cats, dogs, horses, and ruminants, have an additional layer in the choroids called the tapetum lucidum. This causes the animal's eyes to appear to glow when shined with a light. The tapetum Iucidum reflects light back toward the retina so that the animal can see in low light.
In the anterior, the choroid becomes the ciliary body that extends from the ora serrata, the serrated front margin of the retina, to just posterior of the junction of the sclera and cornea. It consists of the ciliary processes and ciliary muscles. The ciliary processes are folds of tissue containing capillaries that secrete aqueous humor. Extending from the ciliary processes to the lens are zonular fibers (suspensory ligaments). The ciliary muscles are a bundle of smooth muscles that alter the shape of the lens in order to allow for near or far vision.
The iris is the colored portion at the front of the eyeball that is shaped like a disc with a hole, the pupil, in the center. The color of the iris is controlled by its number of pigmented cells. A large number of pigmented cells results in a brown color; a low number results in a blue color. The shape of the pupil can vary. It can be round, elliptical, or slitlike. Cats have an elliptical pupil that opens and closes faster than round pupils.
The iris lies between the cornea in front and the lens in back, and is attached to the ciliary processes.
Consisting of circular and radial smooth muscle fibers, the ciliary process regulates the amount of light entering the eye. Parasympathetic signals stimulate the circular muscles to contract during bright light and close vision, causing the pupil to constrict. During dim light or distant vision, sympathetic signals stimulate the radial muscles to contract, causing the pupil to dilate.Retina (the sensory tunic)
The innermost layer of the eye is the retina, which lines the posterior portion of the eyeball. The retina consists of two layers: an outer pigmented layer and an inner neural layer. The pigmented layer is a one- cell-thick layer of melanin-containing epithelial cells, similar to the choroid. These cells also act as phagocytes and store vitamin A.
The neural layer of the retina is multilayered, and grows directly out of the brain during embryonic development. It has three major layers: the photoreceptor layer, the bipolar cell layer, and the ganglion cell layer (Fig. 11.12). The outer and inner synaptic layers separate these layers from each other. Before reaching the photoreceptor layer, light must first pass through the ganglion and bipolar cell layers. Light stimulates the photoreceptors found close to the pigmented layer. This generates signals that travel through the outer synaptic layer to the bipolar cells, and through the inner synaptic layer to the ganglion cells. Interspersed among these cells are two other types of neurons: horizontal cells and amacrine cells. These latter neurons form lateral connections that modify signals along the photoreceptive pathway.
Axons from the ganglion cells collectively form the optic nerve, which exits the eye at the optic disc. Since the optic disc lacks photoreceptors, it is also called the blind spot. The blind spot normally is not apparent since the brain "fills in" information from this area. However, you can demonstrate its presence to yourself by covering your right eye and gazing at the plus sign below.
As you move the position of the book closer or farther from your eye, you will see the large dot disappear:
There are two types of photoreceptors: rods and cones. Rods outnumber cones 20:1—except in birds, which have more cones than rods. Rods have a low light threshold, and therefore are effective in dim light, allowing for the perception of only shades of gray. Cones have a higher threshold for light and provide for color and high acuity vision.
The macula Iutea is an oval region found in the exact center of the posterior of the retina. It contains mostly cones. At its center is a small pit, the central fovea, which contains only cones and where the bipolar and ganglion cells are displaced to the sides. Therefore, light passes unimpeded to the cones. The density of cones in the retina decreases moving from the macula toward the periphery. Since the central fovea has a high concentration of cones, this is the region of the eye with the greatest visual acuity (sharpness of vision). So an animal will focus an object on the fovea to generate greater detail.
The avian retina is avascular, and contains a unique structure called the pecten. This is a black pigmented structure extending from the ventral retina up to just above the area where the optic nerve exits. It is made of blood vessels and pigmented stromal cells, and is thought to serve a nutritive function.
Lens
The lens is a biconvex, transparent, avascular structure that can change its shape in order to focus light on the retina. Located behind the iris, the lens is held in place by the suspensory ligament attaching it to the choroid process.
The lens, enclosed in a thin, elastic capsule, consists of two regions: lens epithelium and lens fibers. The
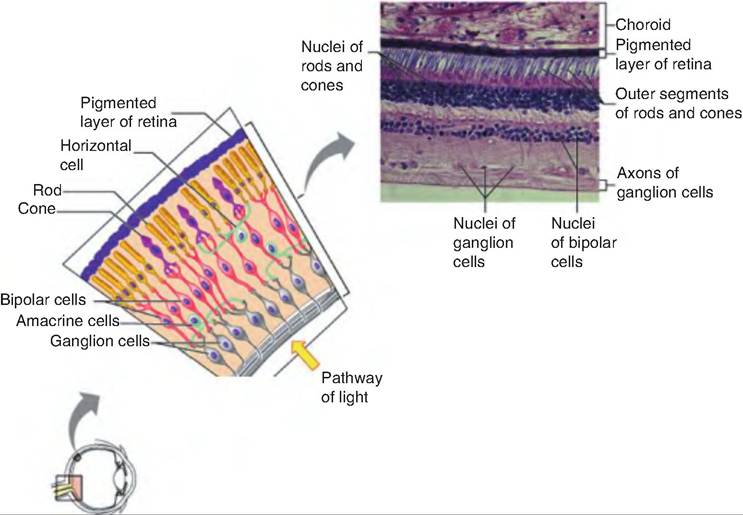
Fig. 11.12. Microscopic anatomy of the retina. Copyright © 2004 Pearson Education. Inc. publishing as Benjamin Cummings.
lens epithelium consists of cuboidal cells located on the anterior surface of the lens. These cells differentiate into the lens fibers that form the bulk of the lens. Arranged like layers of an onion, lens fibers are anu- clear and contain few organelles. Lens fibers are made of folded proteins called crystallins.
Chambers of the eye
The lens divides the eye into the anterior and posterior segments. The iris subdivides the anterior segment into the anterior chamber, located between the cornea and iris, and the posterior chamber, located between the iris and lens. The anterior segment is filled with aqueous humor. It is a clear, watery fluid similar in composition to plasma, which nourishes the lens and cornea. Aqueous humor is continually derived as a filtrate from the capillaries of the ciliary processes entering the posterior chamber. It flows forward through the pupil of the iris into the anterior chamber. From there, it drains into the venous blood via the schleral venous sinus (canal of Schlemm).
Normally produced and removed at the same rate, aqueous humor, along with the vitreous humor discussed below, maintains the intraocular pressure. However, if the drainage of aqueous humor is blocked, intraocular pressure increases, causing compression of the retina and optic nerve. This can lead to glaucoma and blindness.
The posterior segment of the eye is the larger of the two segments. It contains vitreous humor, a clear gellike substance. The vitreous humor pushes the retina against the pigmented layer of the choroid, allowing the retina to receive a clear image. Unlike aqueous humor, the vitreous humor forms during embryonic development and lasts a lifetime. Running through the center of the vitreous humor from the lens to the optic disc is the hyaloid canal, a narrow channel that is occupied by the hyaloid artery during fetal development. Occasionally, debris called vitreous floaters are visible within the vitreous humor. Such debris may cast a shadow on the retina.
Physiology of vision
The eye can be likened to a camera. The image of an object is focused on the retina by the lens, and the amount of light entering the eye is controlled by the pupil. The retina, lens, and pupil are analogous to the film, lens, and aperture of the camera, respectively. Three processes are important in the formation of a clear image: refraction, accommodation, and pupil diameter.
Refraction
When light rays pass from one medium to another of a different density, the speed of light changes. As a result, the light rays are bent, or refracted (Fig. 11.13). For example, when light rays move from air into water, light rays are bent at the interface of the two
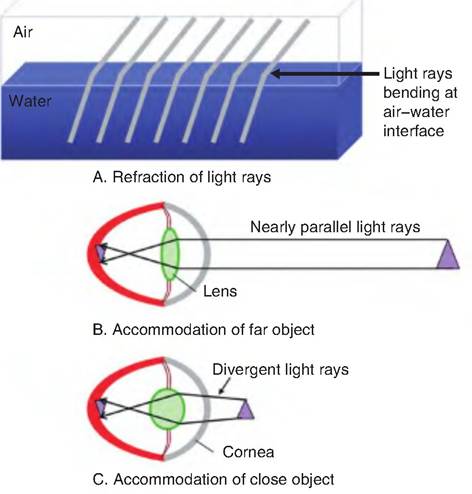
Fig. 11.13. Refraction of light rays. (A) Refraction of light rays at the interphase between air and water. (B) The refraction of light in order to focus a far object on the retina. (C) Note the more spherical shape of the lens necessary to focus a near object on the retina.
mediums. With respect to the eye, light rays are refracted, or bent, at the anterior and posterior surfaces of both the cornea and lens. Approximately 75% of the refraction occurs at the interphase with the cornea. Note that images are inverted, both upside down and backward, as they are focused on the retina. The brain reinterprets this image so that objects are not perceived as inverted.
Accommodation
In the eye, the angle at which the light rays are bent depends on the shape of the lens. The more convex the lens, the greater the degree to which light rays are bent. As an object is moved closer to the lens, the light rays must be bent at a greater angle in order to focus the image on the retina. The process of increasing the refractive power of the lens is called accommodation. Therefore, as an object moves closer to the eye, the lens must become more rounded, that is, made more convex, in order to focus the image on the retina.
Accommodation is accomplished by the actions of the ciliary muscle. When the ciliary muscle is relaxed, the zonular fibers surrounding the lens pull on the lens, thus making it fatter or less convex. When the ciliary muscle contracts, it pulls the ciliary body and choroid forward, thus decreasing the tension of the zonular fibers on the lens. As a result, because of elastic fibers in the lens, it becomes more convex, that is, more rounded, which increases its focusing power, causing greater bending of the light rays.
In addition to the accommodating mechanisms described above, some species of birds possess a static mechanism allowing them to keep objects in focus regardless of their distance. This is accomplished from asymmetries in the eye, allowing it to be emmetropic (i.e., light rays focus directly on the retina) in its upper portions, while becoming increasingly myopic (i.e., nearsighted) toward its lower portions. This allows a bird to keep objects in the horizon in focus in the upper portion of the eye while simultaneously keeping nearer objects in focus in the lower portion of the eye.
Horses appear to have a limited accommodating ability due to weak ciliary muscles. To compensate, horses have a ramp retina in which the distance between the lens and the retina varies from dorsal to ventral positions. This allows horses to use a form of static accommodation in which they move their head to focus an object at different locations on the retina, depending on the distance of the object from the eye.
The far point of vision is that distance beyond which no accommodation (no change in lens shape) is necessary for focusing. The near point of vision is the closest point at which the animal can focus clearly. It is the point of maximum accommodation of the lens. This point gradually gets farther away as an animal ages.
Refraction problems
A normal eye is said to be emmetropic. As animals age, the lens loses its elasticity, and therefore its ability to accommodate, a condition called presbyopia. When an animal can see close objects but distant objects are blurred, it is called myopia or nearsightedness. It occurs because the eyeball is too long relative to its focusing power, and therefore distant objects are focused in front of the retina (Fig. 11.14). In hypermyopia, also called hyperopia or farsightedness, the animal can see distant objects but is unable to focus near objects because the eyeball is too short. Therefore, there is not enough accommodating power to focus the light rays of a near object, and the animal instead focuses at a point behind the retina. An irregular curvature in either the lens or cornea results in astigmatism.
Pupil diameter
The amount of light that can enter the eye is controlled by the diameter of the pupil. Circular muscle fibers control the pupil diameter. During the accommodation papillary reflex, parasympathetic signals from the oculomotor nerves cause the pupil to constrict, thus preventing the most divergent light rays from entering the eye. These light rays would fall on the
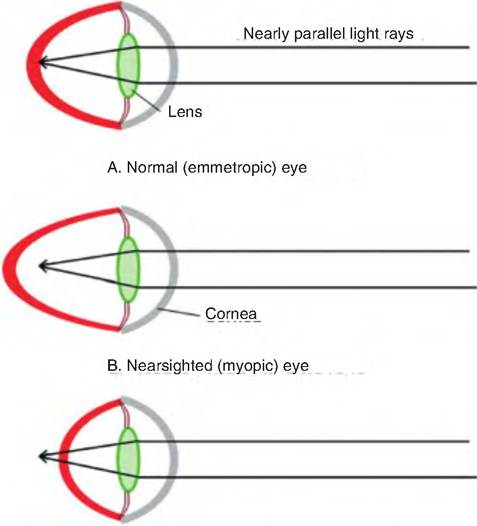
C. Farsighted (hypermytropic) eye
Fig. 11.14. Refraction problems. (A) A normal (emmotropic) eye that can focus light rays on the retina. (B) A nearsighted (myopic) eye is too long; thus, the light rays from distant objects are focused in front of the retina. (C) In a farsighted (Iiypermyopic) eye, the eyeball is too short so that near objects are focused behind the retina, and the lens does not have enough accommodating power to focus the light rays on the retina.
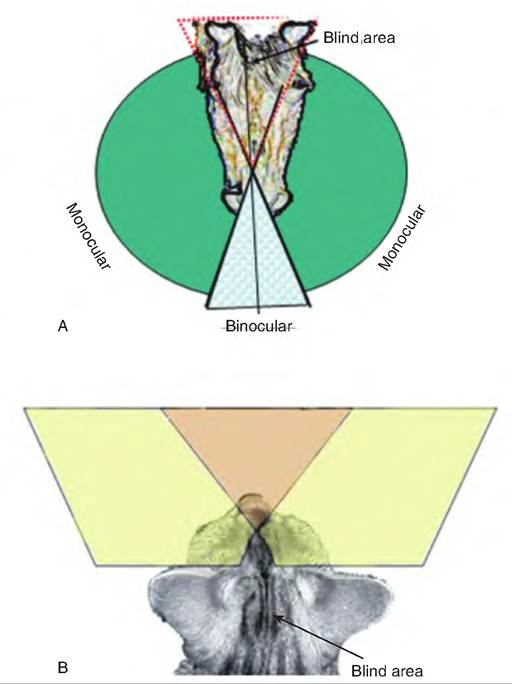
Fig. 11.15. Field of vision. (A) There is a blind area immediately between the eyes of a horse and behind the horse's body. (B) The cat has a larger binocular area than the horse, but also has a greater blind area behind its body.
periphery of the retina where they would not be focused properly.
Field of vision
The field of vision is the spatial area that can be seen by a single eye, providing monocular vision. The location of the eyes within the head has an impact on the field of vision. The field of vision of the two eyes generally overlaps, providing an area of binocular vision. The eye location varies between species and within breeds of species. The wider set the eyes, the greater the panoramic field of vision. Herbivores tend to have their eyes set wide, thus providing them with a panoramic field of vision (Fig. 11.15), although they cannot see directly in front of the nose or behind their hindquarters.
Photoreception
Photoreception involves the light energy focused on the retina being converted to an electrical signal carried by the optic nerve. As mentioned earlier, photoreceptors consist of rods and cones, named for the shape of the outer segment (Fig. 11.16). The outer segment of rods is cylindrical whereas that of the cones is tapered or cone-shaped. The tips of the rods and cones lie next to the pigmented layer. In birds, oil droplets are also found in cones, which may help filter out UV radiation.
Rods and cones consist of an outer segment involved in photoreception and an inner segment containing the cell nucleus, Golgi complex, and mitochondria. The proximal end of each photoreceptor consists of bulb-shaped synaptic endings containing synaptic vesicles.
Within the outer segments are stacks of membranous discs in which the visual pigments, or photopigments, are embedded. In rods, the discs are discontinuous and stacked like pancakes, one on top of the other. In cones, the disc membranes are continuous with the plasma membrane, and the interior of the discs is continuous with the extracellular space. In rods, one to three new discs are added to the base of
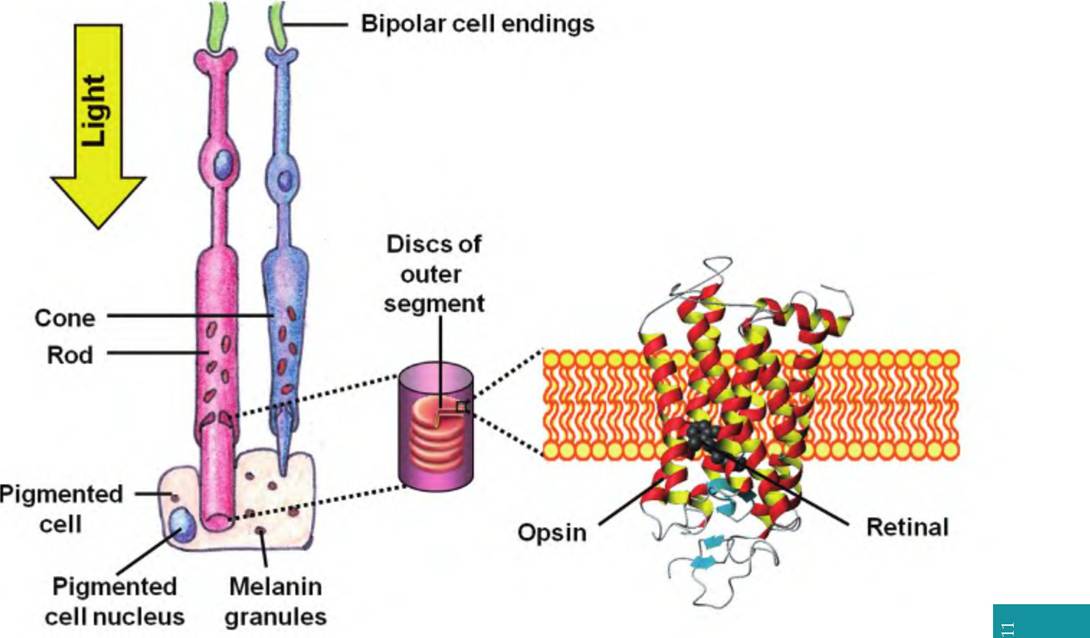
Fig. 11.16. Structure of rods and cones. Rods have a cylindrical outer segment; cones have a conical-shaped outer segment. Note the direction of light and that the tips of the photoreceptors lie near the pigmented layer.
the outer segment every hour, thus pushing old discs toward the distal end where they are sloughed off and phagocytized by the pigmented epithelial cells. Cones also renew their discs, but it appears to occur in a circadian rhythm and is not as well understood.
There is one type of rod and three (four in birds) types of cones, distinguished by different visual pigments. Although rods are more sensitive to light and are stimulated by all visible wavelengths, they perceive only gray tones, whereas cones allow for differentiation of color.
Chemistry of visual pigments
The light-absorbing photopigment in rods is rhodopsin. It consists of a glycoprotein called opsin and a vitamin A derivative called retinal. Vitamin A is found in carotenoid-rich vegetables, including carrots, spinach, broccoli, and yellow squash, as well as vitamin A-containing tissues such as liver. Although retinal is the light-absorbing part of all photopigments, the opsins found in each of the three types of cones differ, permitting them to absorb primarily either blue, green, or yellow-orange wavelengths of light.
The photopigments respond to light in the following sequence (Fig. 11.17):
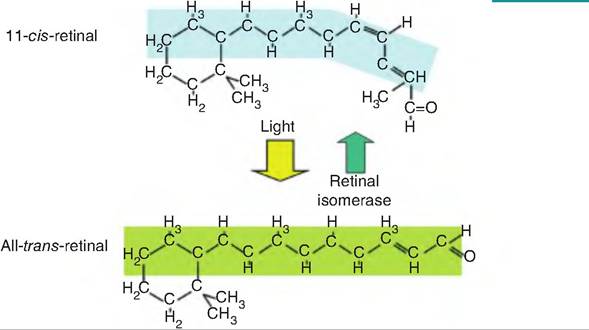
Fig. 11.17. Retinal isomers involved in photoreception. In the dark, 11-c⅛-retinal remains bound to opsin to form rhodopsin. When light strikes the retina, 11-c⅛-retinal is converted to all-trans-retinal in a process called bleaching. All-trans-retinal dissociates from opsin and can be isomerized back to 11-c⅛-retinal in the pigmented epithelium by the enzyme retinal isomerase, which requires ATP.
1. In the dark, retinal is found in a bent configuration called 11-czs-retinal, which binds strongly with opsin. In rods, this forms rhodopsin.
2. When exposed to light, czs-retinal is isomerized to all fraπs-retinal, which is no longer able to bind to retinal. Therefore, fraπs-retinal and opsin dissociate, thereby forming a colorless photopigment. This process is called bleaching.
3. The enzyme retinal isomerase converts trans- retinal back to czs-retinal.
Light transduction by photoreceptors
In the dark, cyclic GMP (cGMP) binds to sodium channels located on the plasma membrane of the outer segment of the photoreceptor and keeps them open (Fig. 11.18). The influx of sodium, called the "dark current," into the photoreceptors depolarizes the membrane. This keeps the Ca2+ channels on the synaptic endings open, thus allowing the continuous release of the neurotransmitter, thought to be glutamate, which then interacts with the bipolar cells. Glutamate induces the production of inhibitozry postsynaptic potentials in bipolar cells, thus inhibiting them.
On the disc membrane, the presence of light converts 11-czs-retinal to all-fra/zs-retinal, causing opsin to dissociate from the photopigment. Opsin then interacts with a G-protein subunit called Iransducin. Trans- ducin activates phosphodiesterase, an enzyme that breaks down cGMP to GMP The breakdown of cGMP causes the sodium channels on the plasma membrane to close. Since sodium entry into the cell decreases while
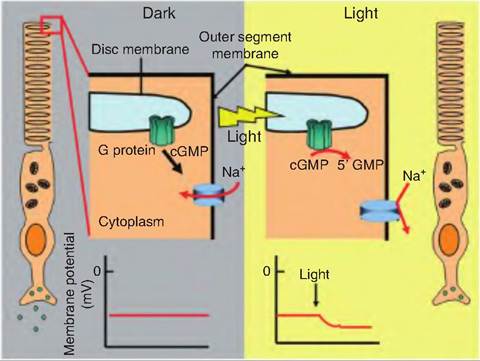
Fig. 11.18. Chemical transduction of light signal. In the dark, cGMP migrates to the plasma membrane and keeps cGMP-gated sodium channels open. When light interacts with rhodopsin found on the disc membrane, it causes 11-c⅛-retinal to be isomerized to all-trans-retinal. Opsin then dissociates from rhodopsin and activates a G protein. The transducin subunit of the G protein activates phosphodiesterate, which breaks down cGMP to GMP. In the light, since cGMP levels are reduced, the sodium channels close, and the cell becomes hyperpolarized, causing a decrease in release of the neurotransmitter glutamate. Since glutamate inhibits bipolar cells, decreased release of glutamate disiinhibits the bipolar cells, so they develop action potential.
potassium leakage from the cell continues, the cell hyperpolarizes as the membrane potential approaches -70 mV, thus decreasing the release of the neurotransmitter. Since the photoreceptors release an inhibitory neurotransmitter, a decreased release of glutamate results in stimulation of bipolar cells. Bipolar cells then stimulate ganglion cells whose axons make up the optic nerve.
Retinal processing of visual information
Only ganglion cells produce action potentials while the amacrine, horizontal, and bipolar cells produce graded potentials. Ganglion cells have circular receptive fields consisting of a circle within a circle (Fig. 11.19). The circular zone at the center is called the on-center area; that on the periphery is called the off- center, or surround, area. The on-center ganglion cells are excited when light illuminates rods or cones in the central area while the off-center area is inhibited. Conversely, light illuminating the off-center area will inhibit the on-center ganglion cells while exciting the off-center ganglion cells.
Each rod or cone in the on-center area synapses in an on-center and off-center bipolar cell (Fig. 11.20). In the dark, rods and cones are depolarized, and glutamate is released. In light, the rods and cones are hyperpolarized, which closes the voltage-gated Ca2+ channels and reduces the release of glutamate. Glutamate affects the on-center and off-center bipolar cells differently. The decreased release of glutamate disin- hibits the on-center bipolar cells, allowing them to become depolarized. Conversely, decreased glutamate release causes the off-center bipolar cells to become
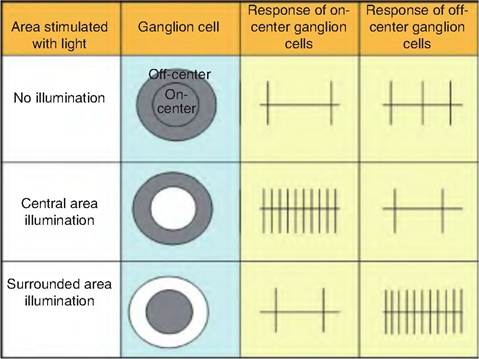
Fig. 11.19. Receptive fields of retinal ganglion cells. Ganglion cells have circular receptive fields with the central area being the on-center area and the surrounding area being the off-center area. On-center ganglion cells increase their firing rate when illuminated; off-center cells have an opposite response, and vice versa when the off-center cells are illuminated.
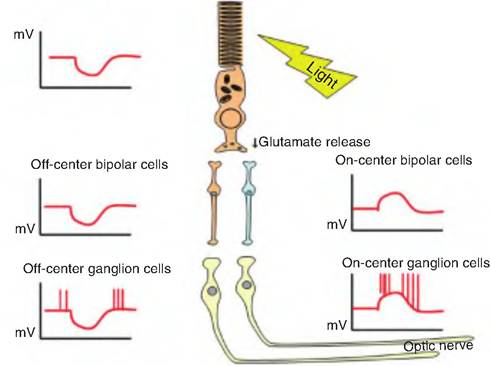
Fig. 11.20. Off-center bipolar cells. The on-center and off-center bipolar cells are differentially affected by glutamate. Glutamate opens Na+-channels on off-center bipolar cells allowing the inward movement of Na+, and thus causing depolarization. Acting on on-center cells, glutamate either opens K+-Channels or closes cGMP-gated Na+-channels, resulting in hyperpolarization. (Modified from Kandel et al., 2000.)
hyperpolarized. The differential effects of glutamate on bipolar cells are due to its effects on different cation channels. On off-center bipolar cells, glutamate opens a Na+-channel allowing Na+ to enter the cell; in on-center bipolar cells, glutamate either opens a K+- channel or closes a cGMP-gated Na+-channel.
Amacrine and horizontal cells function as interneurons providing lateral inputs within the retina. Horizontal cells receive input from the photoreceptors and project to surrounding bipolar cells and photoreceptors, whereas amacrine cells receive input from bipolar cells and project laterally to surrounding ganglion cells, bipolar cells, and other amacrine cells. When the off-center area is illuminated, horizontal cells hyperpolarize the on-center bipolar cells. Similarly, information from surrounding bipolar cells is conveyed to on-center ganglion cells by amacrine cells.
Light or dark adaptation
The eye adapts to a sudden increase in light intensity by decreasing its sensitivity, a process called light adaptation. While light adaptation involves contraction of the pupil to restrict the amount of light reaching the retina and bleaching of photopigments the two most important changes occur in the cone photoreceptors. Bright light causes the cGMP-gated channels to close, thus hyperpolarizing the cells to approximately -70 mV. If bright light continues, the cells slowly depolarize to between -70 and -40 mV so that they are again responsive to light. In addition, cones become desensitized so that the smallest change in light intensity that can be detected by the cell increases with prolonged exposure to light.
Dark adaptation is essentially the reverse of light adaptation. When an animal moves to a relatively dark area, photosensitivity is slowly restored as the rod photopigments regenerate. Cones essentially stop functioning in low-intensity light, so animals can perceive only black, white, and shades of gray. Cats have more rods than cones, making their night vision and motion detection superior to that of humans.
Visual pathway
There are many more rods and cones than bipolar cells, so these photoreceptors converge on bipolar cells, which send signals to the ganglion cells. The axons of the ganglion cells project to the back of the eyeball converging to form the optic nerve. At the optic chiasma, fibers from the medial portion of the eye cross to the opposite side and continue via the optic tracts (Fig. 11.21). Therefore, each optic tract contains fibers from the temporal (lateral) aspect of the eye on the ipsilateral side and fibers from the nasal (medial) aspect of the contralateral eye. Each eye is carrying information from the same half of the visual field from each eye. Since the lens reverses all images, the medial half of each retina receives light rays from the lateral portion of the visual field, while the lateral half of each retina receives light rays from the medial portion of the visual field. Therefore, the right optic tract carries signals representing the left half of the visual field, and the left optic tract carries signals from the right visual field.
The axons of ganglion cells make up the optic nerve, which crosses at the optic chiasma. Most of these axons then travel to the lateral geniculate body of the thalamus, where they synapse on second-order neurons that travel through the internal capsule, forming the optic radiation. These fibers project to the primary visual cortex in the occipital lobes.
Some fibers in the optic tract project to the superior colliculi, which is involved in visual reflexes controlling the extrinsic muscle of the eye. Other fibers project to the pretectal nucleus, which is involved in pupil light reflexes, and to the Suprachiasmatic nucleus of the hypothalamus, which is involved in circadian rhythms.
Hearing and balance
Anatomy of the ear
The ear consists of three regions: the outer, middle, and inner ear. The outer ear, also called the external ear, collects sound waves and directs them to the
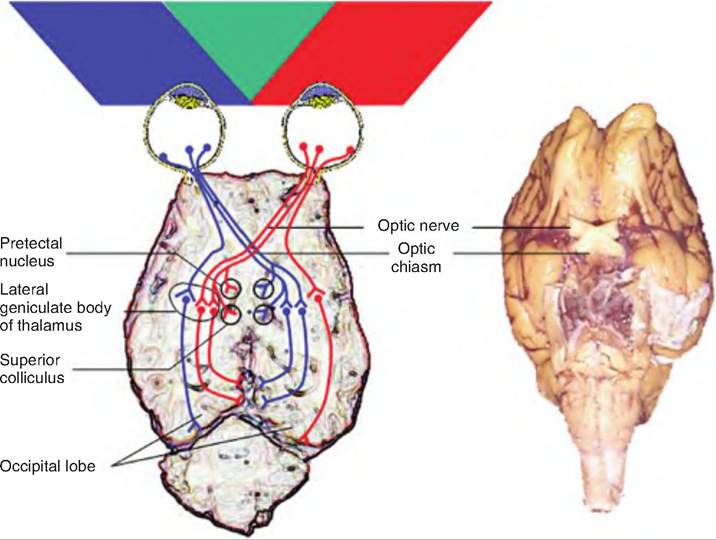
Fig. 11.21. Visual fields of the eyes. The visual fields of the two eyes overlap in the middle, indicated in green. The pathway of signals generated in the retina of the right and left eye are indicated by red and blue lines, respectively.
middle ear. The middle ear carries these sound waves to the inner ear, and the inner ear contains receptors for hearing and equilibrium.
Outer (external) ear
The outer ear consists of the pinna, or auricle, and the external acoustic meatus (Fig. 11.22). The pinna is a fleshy appendage attached to the lateral surfaces of the skull by muscles and ligaments. It is mobile and funnels sound waves toward the external auditory meatus. Hairs and specialized sebaceous (oil) glands called ceruminous glands that secrete earwax are found near the entrance of the external auditory meatus. The hair and earwax function to restrict the entrance of foreign materials such as dust.
Middle ear
Separated from the outer ear by the tympanic membrane, a thin, semitransparent membrane, the middle ear is an air-filled cavity within the temporal bone (Fig. 11.23). It is separated from the inner ear by two openings, the superiorly located oval window and the round window. Extending from the tympanic membrane to the oval window are three auditory ossicles called the malleus, incus, and stapes, or commonly called the hammer, anvil, and stirrup, respectively. These bones are connected by synovial joints. The handle of the malleus is attached to the inner surface of the tympanic membrane, and its head is attached to the body of the incus. The incus articulates with the head of the stapes, whose footplate then fits into the oval windows.
Two small muscles also attach to the auditory ossicles. The tensor tympani muscle restricts the movement and increases tension on the tympanic membrane to minimize damage to the inner ear from loud noises. The stapedius muscle, the smallest skeletal muscle, attenuates large vibrations of the stapes in response to loud noises.
Inner ear
The inner ear is sometimes called the labyrinth because of its extensive series of canals (Fig. 11.23). It consists of two main sections, an outer bony labyrinth enclosing an inner membranous labyrinth. The bony labyrinth lies in the temporal bone and consists of three areas: (1) the semicircular canals, (2) the vestibule, and (3) the cochlea. The first two contain receptors for equilibrium, and the cochlea contains receptors for hearing. The bony labyrinth is lined with periosteum and is filled with perilymph. Perilymph is similar to cerebrospinal fluid. The membranous labyrinth is lined with epithelium and contains endolymph, which contains a relatively high level of K+ ions.
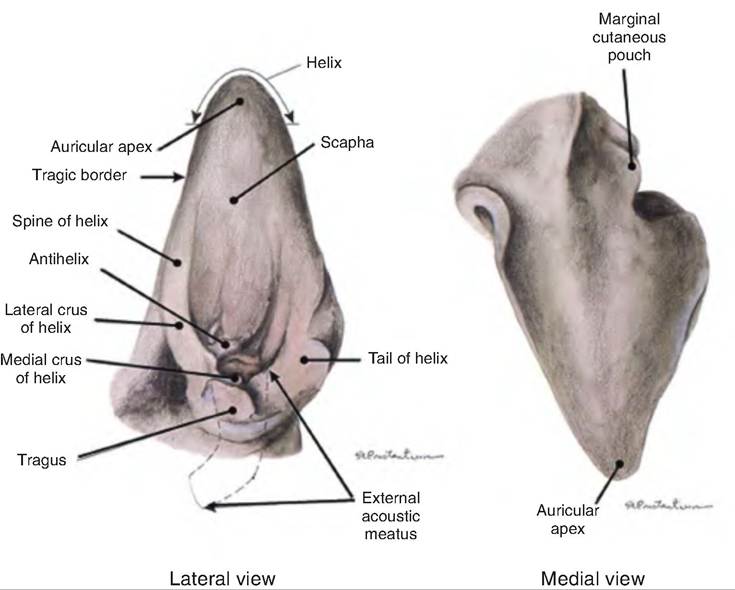
Fig. 11.22. Outer ear of the dog. (Reprinted from Constantinescu, 2002. Used by permission of the publisher.)
The vestibule is the central region of the bony labyrinth. The parts of the membranous labyrinth within the vestibule are the utricle (little bag) and saccule (little sac), which are connected by a short duct. Projecting from the vestibule are three semicircular canals, called the anterior, posterior, and lateral semicircular canals. They lie at a right angle to each other. There is a swollen enlargement at one end of each semicircular canal called the ampulla (saclike duct). The membranous labyrinth within each semicircular canal is called the semicircular ducts, and is continuous with the utricle and saccule.
The cochlea (snail-shaped), a bony spiral canal resembling a snail shell, lies anterior to the vestibule. The cochlea makes nearly three spirals around a bony core called the modiolus. If uncoiled and cut in cross section, the cochlea consists of three channels. Beginning at the oval window, the upper channel is the scala vestibuli. The lower channel is the scala tympani, which ends at the round window. These two channels contain perilymph and are continuous with one another, connecting at the apex of the cochlea, the helicotrema. The third channel, lying between the other two, is the cochlear duct, or scala media. The cochlear duct is separated from the scala vestibuli by the vestibular membrane and from the scala tympani by the basilar membrane.
The spiral organ, or organ of Corti, sits on the basilar membrane. It consists of epithelial cells, supporting cells, and hair cells, the receptors for hearing (Fig. 11.24). There are two groups of hair cells. There is a single row of inner hair cells and three rows of outer hair cells. On the apical (top) surface of each hair cell is a hair bundle consisting of many stereocilia and one long kinocilium, which are in contact with a flexible, gelatinous membrane called the tectorial membrane. At their basal (bottom) end, each hair cell synapses with first-order sensory neurons and with motor neurons from the cochlear branch of the vestibulocochlear nerve (cranial nerve VIII). The cell bodies of these sensory neurons reside in the spiral ganglion. The inner hair cells synapse with approximately 90% of the first-order sensory neurons; 90% of the motor neurons synapse with outer hair cells.
Sound
Sound waves appear as sine waves traveling through a medium such as air or water. Originating from a vibrating object, the pitch is related to the frequency of the sound wave, while the intensity is related to the amplitude of the sound wave. The higher the frequency, the higher the pitch. Similarly, the higher the amplitude, the louder the sound.
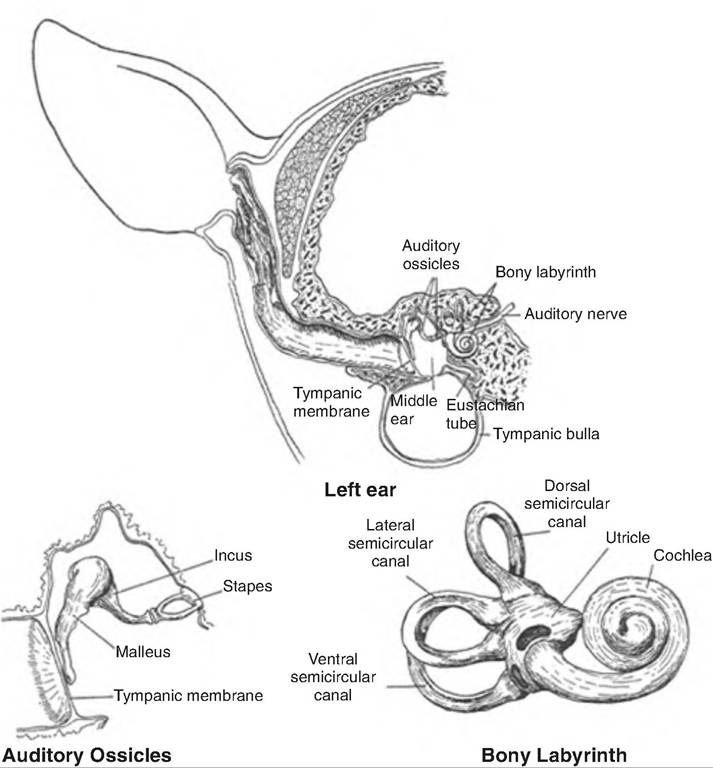
Fig. 11.23. Anatomy of the left ear. (Reprinted from Getty, 1964. Used by permission of the publisher.)
Physiology of hearing
The act of hearing can be described as follows:
1. The auricle collects the sound waves into the external meatus.
2. These sound waves strike the tympanic membrane, causing it to vibrate.
3. The central region of the tympanic membrane makes contact with the malleus, causing it to vibrate. This causes the incus and stapes to vibrate in succession.
4. The stapes makes contact with the oval window, causing it to vibrate. The sound wave is thus transferred to the perilymph in the scala vestibuli located in the cochlea.
5. This sound wave moves through the scala vestibuli, around the helicotrema, and into the scala tympani, which finally causes the round window to vibrate. This sound wave causes the basilar membrane to move up and down.
6. Since the cilia and kinocilia of the hair cells are embedded in the stationary tectorial membrane, as the basilar membrane oscillates, it causes the cilia and kinocilia on the hair cells to shear.
7. If the hair cells shear toward the kinocilia, the hair cell is depolarized, whereas shearing away from the kinocilia causes hyperpolarization.
Depending on the frequency, or pitch, of the sound wave, it causes different regions of the basilar membrane to oscillate. Higher-frequency sounds cause the basilar membrane closest to the round window to vibrate; lower-frequency waves cause the region near the helicotrema to oscillate. The stimulation of hair cells near the round window is associated with high- pitch sounds; those near the helicotrema are associated with low-pitch sounds.
Physiology of equilibrium
In addition to hearing, the ear is involved in the sense of balance. Receptors in the semicircular canals and vestibule collectively make up the vestibular apparatus, the part of the ear associated with equilibrium.
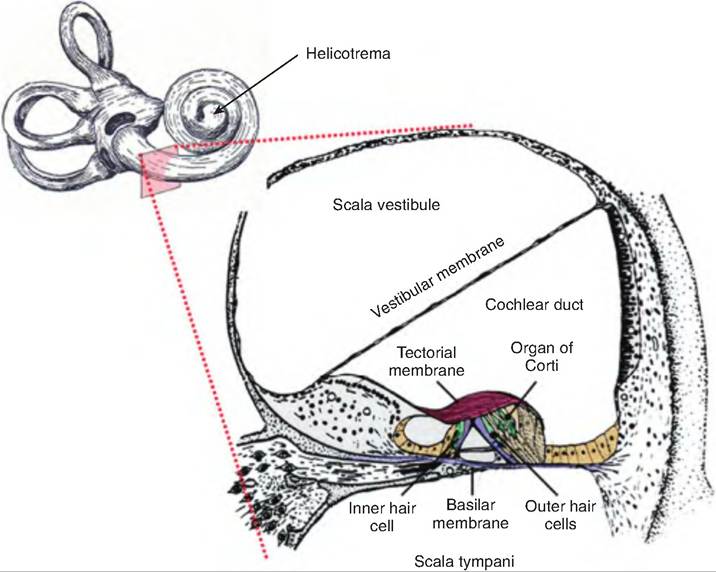
Fig. 11.24. Section through the cochlea. A transverse section through the cochlea reveals its three channels, including the scala vestibule, which is continuous with the scala tympani, and the cochlear duct, which lies in between. The spiral organ (organ of Corti) rests on the basilar membrane. It consists of a single row of inner hair cells and three rows of outer hair cells, which project into the tectorial membrane.
There are two types of equilibrium: static and dynamic. Static equilibrium refers to the maintenance of the position of the body while not moving, whereas dynamic equilibrium is the maintenance of body position in response to sudden movements such as acceleration, deceleration, and rotation.
Otolithic organs within the utricle and saccule
On the walls of both the utricle and saccule is a thickened area called the macula. These are perpendicular to one another and are mostly involved in static equilibrium, but they also have a role in dynamic equilibrium since they respond to linear acceleration, but not to rotation. They provide sensory information about the position of the head in space and thereby function in posture and equilibrium.
Each macula consists of a bed of supporting cells surrounding scattered hair cells. The hair cells have many stereocilia and a single kinocilium projecting from their apex. The hair cells are embedded in an overlying otolithic membrane (Fig. 11.25). The otolithic membrane is a jellylike mass on which crystals of calcium carbonate called otoliths (ear stones) are present. The otoliths provide inertia to the otolithic membrane, which resists any change in motion.
As the head starts or stops moving in a linear direction, the otoliths cause the otolithic membrane to
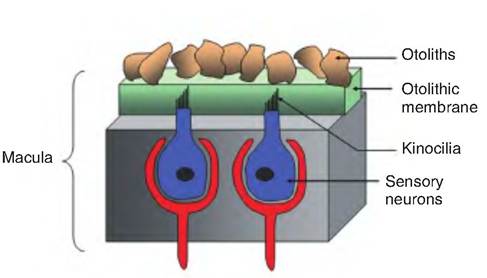
Fig. 11.25. The macula. The macula is a sensory apparatus found in the utricle and saccule. The sensory neurons have hair cells called stereocilia that project into the gelatinous otolithic membrane. The longest stereocilia is called the kinocilium. Embedded in the otolithic membrane are grains of calcium carbonate called otoliths.
shear in the opposite direction, much like the head falls backward when one suddenly accelerates forward in a car. This shearing of the otolithic membrane causes the hair cells projecting into the membrane to bend in the opposite direction of movement. When the hair cells bend toward the single kinocilium, they depolarize; bending in the opposite direction causes hyperpolarization. While the hair cells continuously release neurotransmitters, a change in membrane potential alters the release of the neurotransmitter released by these receptors. This changes the rate of impulse in the vestibular nerve, a subdivision of cranial nerve VIII.
Remember, the macula responds only to changes in linear acceleration of the head. The hair cells adapt quickly, so they do not respond while the head is stationary.
The crista ampularis and dynamic equilibrium
The crista ampularis, located in the ampulla of each semicircular canal, is the receptor for dynamic equilibrium. Like the macula, the crista ampularis consists of supporting cells and hair cells. The hair cells have stereocilia and one kinocilium that project into a gelatinous mass called the capula.
The crista ampularis responds to changes in velocity of rotation of the head. The inertia exerted by the endolymph in the semicircular canals causes the hair cells to bend in the opposite direction of movement. As in the macula, this causes either depolarization or hyperpolarization of the hair cells, thus changing the release of the neurotransmitter.
Signals from the semicircular canals are important in vestibular nystagmus, a reflex movement of the eyes. As the head rotates, the eyes drift in the opposite direction as if fixed on an object. After a time, the eyes rapidly jump toward the direction of rotation and establish a new fixation point. This type of eye movement continues until the endolymph comes to rest.