Gustation: The sense of taste
Tongue
The tongue is the muscular organ filling most of the oral cavity. It is composed of interlacing bundles of skeletal muscle fibers, and it is involved in gripping, repositioning food, mixing food with saliva, and forming the compact mass of food called a bolus.
The tongue has intrinsic and extrinsic muscles. The intrinsic muscles confined to the tongue, and not attached to bone, run in several directions, allowing the tongue to change shape as necessary for prehension, moving food, and making sounds. The extrinsic muscles attach the tongue to bones of the skull and the soft palate. They allow the tongue to protrude, retract, and move side to side. The lingual frenulum attaches the tongue to the floor of the mouth.
The surface of the tongue is covered with small bumps called papillae (Latin for "bumps"; singular = papilla). Papillae are named for their shape. Filiform papillae are thorn-shaped, giving the tongue roughness and thus aiding in licking and manipulating food. They have a mechanical function. In the ox and cat, they are heavily cornified. Fungiform papillae are mushroom-shaped, scattered among the more numerous filiform papillae, have taste buds, and are thus mechanical and gustatory (Fig. 11.2). Foliate papillae have a series of leaf-shaped ridges, are located on the lateral borders of the tongue, and have a gustatory function. They are absent in the ox. Vallate, or circumvallate, papillae are the largest and least numerous. They are located in a V-shaped row near the back of the tongue. They resemble the fungiform papillae, but are circled by a cleft containing taste buds. Marginal taste buds are found along the edge of the rostral portion of the tongue of newborn dogs, but they disappear when puppies switch to solid food.
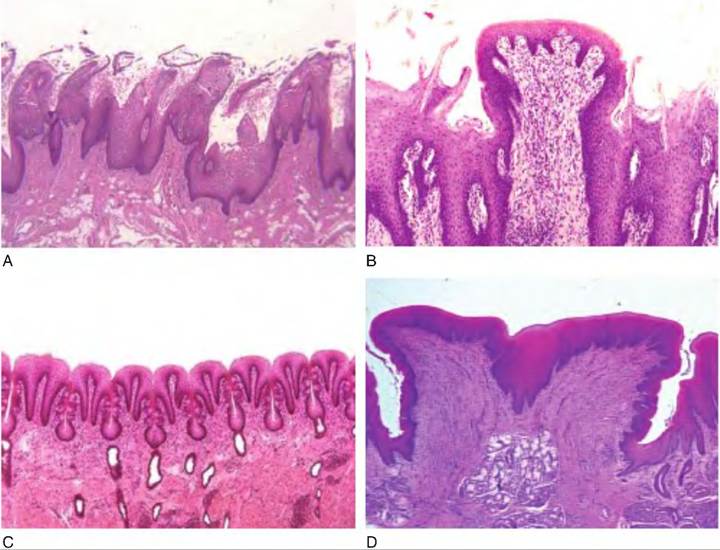
Fig.
11.2. Shape of papillae. (A) Filiform papillae, which are thorn-shaped. (B) Fungiform papillae, which are mushroom-shaped. (C) Filiate papillae, which are leaf-shaped. (D) Vallate, or Circumvallae papillae, which are similarly shaped to fungiform, but have clefts along the side that contain taste buds.Taste
Taste is an important sense in animals because it helps differentiate nutritious feedstuffs from toxic materials. Animals have innate preferences for certain flavors, such as those that are sweet, while tending to reject those that are unpleasant, such as bitter flavors. Each papilla has one to hundreds of taste buds. Taste buds are also located in the palate, pharynx, epiglottis, and upper third of the esophagus.
Each taste bud is gourd-shaped and consists of three types of taste cells, called light, intermediate, and dark cells, as well as basal cells (Fig. 11.3). The different taste cells are believed to be either cells at different stages of differentiation, with the lightest being the most mature, or cells of different cell lineages. The basal cells are thought to be stem cells for the taste cells, which are short-lived. There is a small opening at the surface of the epithelium called the taste pore. Each taste cell has microvilli extending into the taste pore. The microvilli are the only part of the cell exposed to the oral cavity. Each taste cell is innervated by sensory gustatory neurons. The space between the taste cells and sensory neurons appears to act as a synapse. Taste cells are electrically excitable and have voltage-gated Na+, K+, and Ca++ channels on their surface.
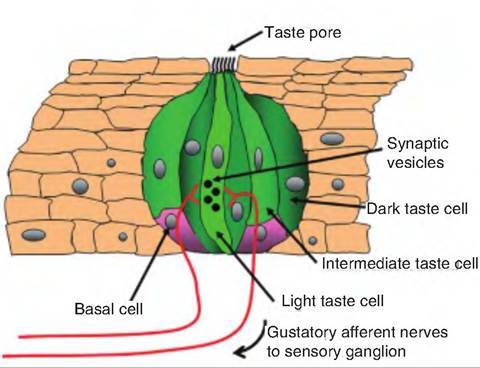
Fig. 11.3. Taste bud. Each taste bud consists of three types of taste cells called dark, intermediate, and light cells. In addition, there are small, round basal cells located at the base of the taste bud. The basal cells are thought to be stem cells that produce taste cells.
Until recently, it was generally believed that animals had four primary tastes, including salty, sour, sweet, and bitter. Recently, a fifth category was added, called umami, meaning "delicious" in Japanese. It is stimulated by monosodium glutamate. Foods can provide a different taste by stimulating a varying combination of these five tastes. In addition, volatile components of feedstuffs stimulate the sense of smell that influences the sense of taste.
The binding of taste stimuli, called tastants, to extracellular receptors located on taste cells leads to depolarization of the taste cell either directly or indirectly through the activation of second messengers. The receptor potential induced by depolarization allows the influx of Ca++ through voltage-gated Ca++ channels. This leads to the release of a neurotransmitter, whose identity is unknown, at the synapse with the sensory gustatory neuron. This neurotransmitter causes excitation in the sensory neuron.
Sweet taste
Sweet taste is initiated by the binding of sweet tastants to receptors. There are two mechanisms for transducing sweet taste stimuli (Fig. 11.4). In the first, the binding of sweet tastants to specific receptors activates a G protein, called gustducin, which activates adenylyl cyclase, causing the formation of cAMP in the cytoplasm. The cAMP then activates protein kinase A, which phosphorylates K+-Selective ion channels located on the basolateral membrane of the taste cells, causing them to close. The cAMP-dependent closure of K+ channels results in depolarization of the taste cells, leading to the release of the neurotransmitter that excites the sensory neuron. In the second mechanism, sweet tastants cause production of G proteins, leading
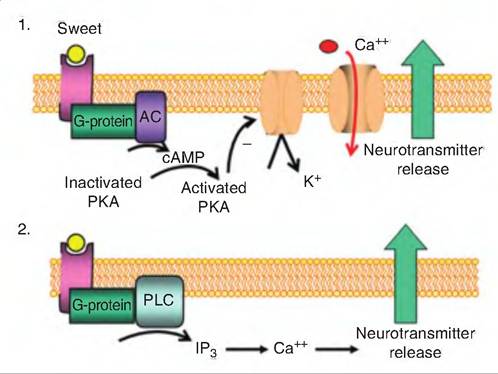
Fig. 11.4. Sweet taste. There are two mechanisms for sweet taste.
(1) A sweet tastant binds to a specific receptor inducing the formation of a G protein that activates adenylyl cyclase (AC), thus causing the formation of cAMP.
The cAMP activates protein kinase A (PKA) which then closes membrane-bound K+-channels, thus causing depolarization of the taste cell. This allows voltage-gated Ca channels to open, and Ca enters the cell, causing the release of the neurotransmitter that excites the gustatory sensory neurons.(2) G proteins activate phospholipase C (PLC), which catalyzes the formation of 1,4,5-triphosphate (IP3). IP3 causes the release of calcium from endogenous stores in the endoplasmic reticulum. Ca then causes the release of the neurotransmitter.
to activation of phospholipase C, an enzyme that catalyzes the synthesis of inositol 1,4,5-trisphosphate (IP3). IP3 causes the release of Ca++ from intracellular stores. Calcium then causes the release of the neurotransmitter, which then stimulates the gustatory sensory neurons.
Bitter taste
Bitter taste, often associated with toxic compounds, is elicited by divalent cations, some amino acids, and alkaloids. Some bitter-tasting compounds (such as quinine) are membrane permeable, while others (such as denatorium) are not, suggesting multiple mechanisms for transduction. The mechanisms for bitter taste appear similar to those for sweet tastes (Fig. 11.5). Denatorium causes the increase in intracellular calcium through an IP3-dependent mechanism. Other bitter compounds activate gustducin that stimulates taste cell phosphodiesterase reducing intracellular cAMP and cGMP levels. Knockout mice, in which the gustducin gene has been deleted, are insensitive to selected bitter tastes. Some bitter compounds are also thought to bind directly to K+-Selective ion channels blocking them. This prevents the movement of K+ out of the cell, thus causing depolarization and release of the neurotransmitter.
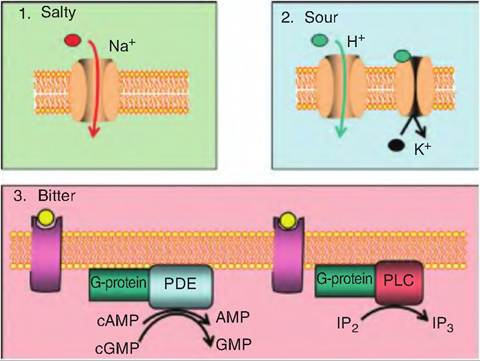
Fig. 11.5. Salty, sour, and bitter taste.
(1) Salty taste is elicited by cations, primarily Na+, which enter through a Na+-Selective ion channel that can be blocked by amiloride. The entry of Na+ causes depolarization of the taste cell and release of the neurotransmitter, which causes excitation in the gustatory neurons. (2) Sour taste is caused by the entry of H+ ions that permeate amiloride-sensitive Na+ channels; this leads to depolarization of the cell because the cell cannot distinguish between Na+ and H+. Sour taste can also be elicited by H+ binding to and blocking K+-Selective channels, preventing the movement of K+ out of the cell. (3) Bitter taste is caused by either increasing intracellular calcium through an IP3-dependent mechanism, or by activating the G-protein gustducin that stimulates taste cell phosphodiesterase (PDE), thus reducing intracellular cAMP and cGMP levels.Salty taste
Salty taste is mostly caused by the cation Na+. Taste cells sensitive to salt have a Na+-Selective ion channel that can be blocked with amiloride (Fig. 11.5). This amiloride-sensitive channel is different from the voltage-gated sodium channels responsible for the upstroke of the action potential. Instead, this channel is not voltage sensitive, and remains open all the time. When an animal consumes feedstuffs high in sodium, this increases the Na+ concentration outside the saltsensitive taste cell, and Na+ then moves down its concentration gradient into the cell, causing depolarization.
Sour taste
The sour taste is caused by the acidity of the feedstuff. The higher the H+ ion concentration, the more sour the flavor. The H+ ions can affect the sour taste receptors in two ways. First, H+ can permeate amiloride-sensitive Na+ channels, which leads to depolarization of the cell since the cell cannot distinguish between Na+ and H+.
Second, H+ can bind to and block K+-Selective channels, preventing the movement of K+ out of the cell (Fig 11.5).Umami
Since amino acids are a major part of dietary intake, it should come as no surprise that animals have a taste for amino acids. Studies with glutamate and aspartate show at least two pathways for this taste. Glutamate can activate an ion channel, allowing Na+ and Ca++ to enter the cell (Fig. 11.6). This inward current causes
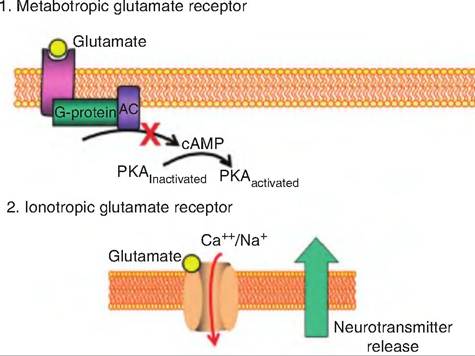
Fig. 11.6. Umami taste. There are two proposed umami pathways: (1) The first involves a glutamate binding to a receptor which activates a G protein (i.e., a metabotropic receptor). The activated G protein causes a decrease in cAMP levels, resulting in a decrease in the activation of phosphokinase A (PKA), and thus a decrease in the metabolic pathway downstream. (2) The second pathway involves an ionotropic receptor in which glutamate interacts with a receptor on the taste bud surface causing the influx of Na+ and Ca++. The resulting depolarization results in neurotransmitter release.
voltage-gated Ca++ channels to open, causing depolarization that triggers neurotransmitter release. The second mechanism entails glutamate binding to a G protein-coupled receptor, probably resulting in a decrease in intracellular cAMP levels.
Taste pathway
Information from taste buds in the anterior two-thirds of the tongue and on the palate travel in the chorda tympani, a branch of the facial nerve (cranial nerve VII). Signals from the taste buds in the posterior third of the tongue are carried via the glossopharyngeal nerve (IX). Taste buds on the epiglottis and esophagus are innervated by the vagus (cranial nerve X). Fibers carrying this information enter the solitary tract in the medulla and enter the gustatory area of the nucleus of the solitary tract. Second-order fibers then ascend to the thalamus, synapsing in the parvocellular region of the ventral posterior medial nucleus. This area then projects to the area along the border between the anterior insula and frontal operculum in the ipsilateral cerebral cortex. Note that this pathway does not cross to the contralateral side.