WEST NILE VIRUS INFECTION
KAROLY ERD ELYI
Central Agriculture Office, Veterinary Diagnostic Directorate, Budapest, Hungary
West Nile virus (WNV) infection (also known as West Nile fever and West Nile disease) infects a wide range of vertebrate species, and its clinical manifestations range from asymptomatic infection, through febrile and neurological disease to acute fatal infection, depending largely on the affected species and the virulence of the virus strain.
AETIOLOGY
WNV belongs to the Flavivirus genus in the Flaviviridae family. From the 12 viral sero- complexes defined within the Flavivirus, WNV is assigned to the Japanese encephalitis virus complex. This group comprises genetically and antigenically closely related viruses such as the Japanese encephalitis virus, Murray Valley encephalitis virus, St Louis encephalitis virus, Usutu virus, WNV and others.
WNV was isolated for the first time in 1937 in Uganda from a febrile human patient. This virus has a wide distribution, extending from Africa and Madagascar over Southern and Eastern Europe to the Middle East, India, Central Asia and Australia. The virus was reported for the first time in the western hemisphere in 1999(1) and has subsequently extended its distribution there.
WNV strains can be classified into at least six separate lineages, which significantly differ from each other, exhibiting wide genomic diversity (76—77% nucleotide iden- tity)(2). Lineage 1 WNV strains can be divided into three clades, which are also related to the geographical distribution of these viruses. Clade 1a comprises European, African and, since 1999, North American strains, whereas Australian (Kunjin virus) and Indian isolates form clade 1b and 1c, respectively. Lineage 2 WNV strains had been mostly isolated in southern Africa and Madagascar, but their endemic presence has been recorded in Central- Eastern Europe since 2004(2).
Rabensburg virus strains establishing lineage 3 of WNV were isolated in the Czech Republic from Culexpipiens mosquitoes, whereas lineage 4 is represented by a single virus (LEIV- Krnd88- 190) isolated in 1998 in Russia. Recently analyzed WNV strains isolated in India during the second half of the 20th century form an additional lineage 5 of WNV.WNV is an enveloped virus with a small virion of about 40-50 nm in diameter. The round nucleocapsid shows icosahedral symmetry, and it is surrounded by a host- derived lipid envelope. The complex nucleocapsid structure consists of the viral RNA and multiple capsid protein molecules. The WNV genome is an 11,000-base-long, linear, positive-sense, single-stranded RNA strand, lacking the polyadenylated tail at the 3' terminus but containing a methylated nucleotide cap at the 5' end of the genome and several conserved secondary motifs regulating viral replication. The cross- neutralization analyses and monoclonal antibody binding assays reflect the genomic differences and antigenic variability among virus strains(3).
Epidemiology
GEOGRAPHICAL DISTRIBUTION IN EUROPE
Evidence of WNV infection in Europe was first detected in 1958, when WNV- seropositive human patients were found in Albania. In Europe the virus was simultaneously isolated for the first time in 1963 from humans and mosquitoes from the Rhone delta, France, and from humans and Hyalomma marginatum ticks from the Volga delta, Russia. Further WNV isolates were obtained soon after from humans, wild birds, horses, rodents, mosquitoes and ticks in Portugal, Slovakia, Moldavia, Ukraine, Hungary, Romania, Czechoslovakia, Italy and other countries(4,5). Early epidemiology was based on the study of WNV epidemics in Egypt during the 1950s. Human West Nile fever cases were diagnosed from the 1960s in the South of France, Russia, Spain, Romania, Belarus, Ukraine and Czechoslovakia1-4). A new wave of outbreaks, generally associated with significant human morbidity and mortality, emerged in the 1990s in Algeria, Romania, Tunisia, Russia,
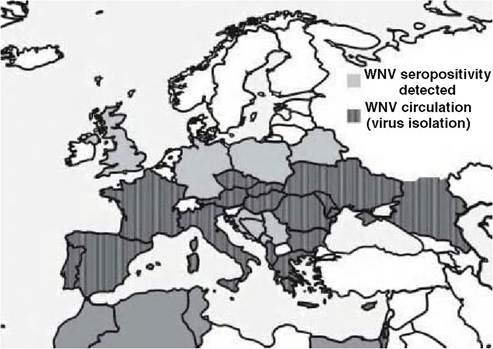
FIGURE 9.1 Map of Europe, indicating the areas of WNV presence and circulation (dark grey) and countries where seropositivity (pale grey) to WNV has been detected in birds or mammals.
Israel and Sudan. During the same period, WNV outbreaks in the Camargue region of France and in Northern Italy affected primarily horses, but human cases were also recorded. In 2003 an outbreak of lineage 1 WNV affected a flock of domestic geese in the South of Hungary. The first occurrence of a lineage 2 WNV strain on the European mainland was detected in infected birds of prey in the southeast of Hungary during the summer of 2004(6). This viral strain has become enzootic in the Carpathian Basin, reaching Austria in 2008 on a westward range expansion and causing an unexpected human outbreak in Greece in 2010. Lineage 1 viruses continued to circulate in Northern Italy during 2008-2009, affecting primarily horses and humans. WNV lineage 1 activity was also detected on the Iberian Peninsula between 2001 and 2007 in both Spain and Portugal. Figure 9.1 shows areas of WNV presence and circulation and countries where seropositivity to WNV has been detected in birds or mammals in Europe.
HOST FACTORS
In contrast to the high rates of clinical disease and mortality reported in North American wild bird species following the 1999 introduction and subsequent expansion of WNV in the western hemisphere, the circulation of WNV in the Old World has not resulted in mass mortality or disease outbreaks in native European bird species. However, closer monitoring and targeted examinations recently revealed that some species regularly succumb to WNV infection in Europe. WNV-related clinical disease and mortality in both wild and captive birds of prey has been regularly demonstrated in Hungary since 2004. Wild and captive (falconry) goshawks (Accipitergentilis) are most often diagnosed with the disease, frequently with high mortality (e.g. 27 WNV- r elated mortality cases in Hungary were confirmed by both pathology and virology between 2004 and 2009). Non-native gyr falcons (Falco rusticolus) and Harris hawks (Parabuteo unicinctus) and a single native peregrine falcon (Falco peregrinus) kept for falconry also suffered WNV-related mortality in the area.
WNV infection with mortality was diagnosed in nestlings and fledglings of free ranging red footed falcons (Falco vespertinus) and sparrowhawks (Accipiter nisus) in Hungary. Goshawks and a captive bearded vulture (Gypaetus barbatus), snowy owl (Bubo scandiacus) and keas (Nestor notabilis) succumbed to WNV infection during the Austrian expansion of lineage 2 WNV. In Spain, WNV-associated disease and mortality was diagnosed in Spanish imperial eagles (Aquila adalbertι) between 2001 and 2005(7) and a golden eagle (Aquila chrysaetos) in 2007(8). Further cases of non- fatal clinical WNV infections were diagnosed in a golden eagle and a Bonelli’s eagle (Hieraaetus fasciatus) in 2007. The virus strains isolated from these birds belonged to lineage 1 of WNV. Mortality of WNV-infected subadult white storks (Ciconia ciconia) was recorded in Israel during the autumn migration in 1999(9).Clinically healthy birds were infected with lineage 1 WNV during the 2008 WNV epidemic in Northern Italy. WNV was demonstrated by reverse transcription polymerase chain reaction (RT-PCR) in samples collected from magpies (Pica pica), carrion crows (Corvus corone), rock pigeons (Columba livia), Eurasian jays (Garrulus glandar- ius), starlings (Sturnus vulgaris), one cormorant (Phalacoc- orax carbo^) and a yellow-legged gull (Larus michahellis). WNV belonging to lineage 1 was also isolated in 2004 from a magpie and a house sparrow (Passer domesticus) found dead in the Camargue, France. A single magpie shot in the area of the 2010 Greek epidemic was found infected with the lineage 2 WNV strain also isolated from the human cases and mosquitoes. Additional WNV isolates were also obtained earlier from the following wild bird species in Europe: barred warbler (Sylvia nisoria), turtle dove (Streptopelia turtur), pied wagtail (Motacilla alba) on autumn migration, and green sandpiper ( Tringa ochropus), lapwing (Vanellus vanellus), black-headed gull (Larus rid- ibundus) and turtle dove returning on migration from their wintering grounds.
Serological studies detected prior exposure to WNV in: house sparrows, magpies, scops owls (Otus scops), willow warblers (Phylloscopus trochilus), whitethroats (Sylvia communis), woodchat shrikes (Laniussenator), hoopoes ( Upupa epops), blackcaps (Sylvia atricapilla), common redstarts (Phoenicurus phoenicurus) and pied flycatchers (Ficedula hypoleuca) in the Camargue, France; in common coots (Fulica atra) in Spain; in pied flycatchers (Ficedula hypoleuca), common redstarts (Phoenicurus phoenicurus), red- backed shrikes (Lanius collurio), garden warblers (Sylvia borin), white storks (Ciconia ciconia), ospreys (Pandion haliaetus), red kites (Milvus migrant), a goshawk and mute swans ( Cygnus olor) in Germany; in sparrows, white stork nestlings, mute swans and a hooded crow ( Corvus corone) in Poland; in common coots, common kingfishers (Alcedo atthis), reed warblers (Acrocephalus scirpaceus), sedge warblers (A. schoenobaenus), marsh warblers (A. palustris), Savi’s warblers (Locustella luscinioides), reed buntings (Emberiza schoeniclus), blackcaps, penduline tits (Remiz pendulinus), blue tits (Parus caeruleus) and starlings in the Czech republic; and other wild bird species in the UK, Sweden, Spain, Portugal, Belarus, Bulgaria and Romania.
Humans and horses are the only mammalian hosts in which clinical WNV is reported in European endemic areas. WNV has occasionally been isolated from small mammal species (e.g. bank voles ( Clethrionomys glareolus), yellow-necked mice (Apodemus flavicollis), domestic mammals (e.g. sheep, cattle) and amphibians (lake frog (Rana ridibunda)). Seropositivity to WNV was also detected in a number of domestic and wild mammalian species(4’10).
ENVIRONMENTAL FACTORS
West Nile fever in Europe occurs primarily during the period of peak mosquito activity (July to September). Environmental determinants of mosquito activity and population dynamics are crucial for triggering WNV outbreaks. Mosquitoes are the primary vectors involved in the transmission of WNV, and the virus has been detected in at least eight mosquito species in Europe and more than 60 species in North America.
In Europe, the virus has been isolated from Culex modestus, C. pipiens, Mansonia richar- di(, Aedes cantans, A. caspius, A. excrucians, A. vexans and Anopheles maculipennis. Culex pipiens is the most significant vector species among the above because of its preference for feeding on birds. The potential to serve as a true vector of WNV depends on several characteristics, such as biological competence, host preference and population density. It has been suggested that, depending also on the genotype of the virus, higher ambient temperatures may effectively shorten the WNV transmission cycle and thus also accelerate the geographic spread of the disease. WNV may spill over from its endemic bird—mosquito cycle through ‘bridge vectors’ that feed on both bird and mammal hosts. A seasonal shift towards preference for mammal hosts later in the transmission season has been observed in C. pipiens and C. tarsalis, which may enhance their importance as bridge vectors. Although WNV has been detected in several tick species (i.e. Hyalomma marginatum, Dermacentor marginatus in Europe) the competence of soft or hard ticks as true vectors of WNV is still uncertain. It has been indicated that overwintering infected vectors also play a role in the maintenance of endemic WNV cycles under different climatic conditions.Epidemiological role of
THE AFFECTED SPECIES
Birds are the principal hosts of WNV, and viraemic birds maintain the infectious cycle through ornithophilic mosquito vectors. It is not yet known which European bird species may serve as reservoirs, amplifying or transporting hosts of WNV. The repeated WNV outbreaks in Europe indicate that the virus is regularly (re)introduced from endemic areas in Africa by infected migratory birds. Migratory passerines have been identified as potential longdistance carriers of WNV in North America, but their potential to play a similar role in Europe has not been proven. Larger trans- Saharan migrant species (e.g. white storks) are probable candidates for this role. The monitoring of the local amplification of WNV and the short distance dispersal of WNV has been attempted in abundant, relatively sedentary species such as magpies, house sparrows and feral pigeons. Preliminary results indicate that these species could be suitable indicators of local WNV circulation, but their role in the epidemiology of the disease is, as yet, unknown.
The level of viraemia in mammals is generally considered to be too low to allow for a significant role of mammalian hosts in the maintenance of the natural cycle of WNV, and mammals are usually considered to be deadend hosts. However, experimental evidence from several North American small mammals (e.g. fox squirrels ( Sciurus niger), eastern grey squirrels (S. griseus), western grey squirrels (S. carolinensis), eastern chipmunks (Tamias striatus) and eastern cottontail rabbits (Sylvilagus floridanus)) suggests that some mammalian species may play a role in the maintenance of the natural WNV cycle.
Studies during the WNV epidemic in North America revealed that a wide range of wild and domestic mammals as well as reptiles (over 30 species) acquire the infection and seroconvert; furthermore, some species may develop high levels of viraemia. The isolation ofWNV strains from wild rodents in Hungary during the 1970s also points to the potential role of small mammals in the maintenance of endemic WNV foci.
TRANSMISSION
Although WNV is predominantly transmitted by mosquito bites in a bird—mosquito cycle, there are also alternative routes of WNV transmission. The spread of WNV by the oral route has been suggested by both experimental results and the circumstances of certain natural WNV infection cases of scavengers and birds of prey in both North America and Europe. Circumstantial evidence for this hypothesis is derived from the explosive geographic expansion ofWNV (beyond the speed modelled for a pure mosquito-borne cycle) and the high incidence of WNV in scavenging corvids, and in diurnal and nocturnal birds of prey in North America, as well as in goshawks in Hungary and Austria. It has been found that infected birds may excrete significant quantities of the virus in their faeces (>108pfu∕g in American crows), so the potential importance of the horizontal faecal-oral transmission route has still to be fully assessed(3).
PATHOGENESIS, PATHOLOGY AND IMMUNITY
The primary route of WNV transmission is through the bite of infected mosquitoes. A viraemia in the bitten animal or bird, of varying length and intensity, ensues after an initial stage of localized viral replication at the inoculation site and in regional lymph nodes. Birds usually develop high levels of viraemia, which may last for 7—8 days. The viraemic stage ofWNV infection in mammals is short and circulating virus levels are generally low. The virus is disseminated to its primary targets, the brain, meninges, spinal cord, heart, kidneys and spleen, as well as other organs. WNV replicates in a range of cell types but with a high preference for neurons, for example Purkinje cells, myocardiocytes, tissue macrophages, blood monocytes, renal cells and intestinal crypt epithelial cells(11). Virus replication within the target cell results in cell degeneration and necrosis, accompanied by focal and perivascular or often extensive, diffuse inflammatory reactions.
I n birds, antibodies to WNV appear 7—11 days postinfection (pi) and may persist for more than a year. Passive immunity transmitted vertically to offspring by antibody transfer into the yolk of bird eggs was found to last on average for 27 days in pigeon squabs. The clearance of WNV infection is achieved primarily through the effect of neutralizing antibodies, but both humoral and cellular mechanisms play a role in modulating immune reactions to WNV. Genetic susceptibility of the host species, host age and immune status, virus strain, route of infection and the infectious dose may all influence the pathogenesis of WNV infection(3).
Macroscopic lesions are non-specific. The usual findings are signs of circulatory collapse, general congestion of internal organs and cardiac enlargement. Affected birds and mammals may be in poor body condition. More sporadic findings, including a pale, mottled myocardium, splenomegaly and leptomeningeal haemorrhages, may also be associated with WNV infection.
Microscopic lesions associated with WNV infection are almost always present in the brain and the heart(6). Severely affected birds usually have a multifocal, lymphocytic panencephalitis accompanied by marked gliosis, and neuronal degeneration comprising chromatolysis, necrosis and neu- ronophagia. Lesions in the brain are frequent in the outer layer of the cerebellum. Multifocal haemorrhage can be present in some cases and a predominantly lymphocytic meningitis may also be a prominent feature. Perivascular cuffing by lymphocytes and plasma cells may be irregular or incomplete. Lesions in the cerebellum and brainstem are often mild. Spinal cord lesions are usually found within the grey matter and focal mononuclear infiltrations can also be present in the peripheral nerves of the sciatic plexus. Degenerative changes detected in association with inflammatory lesions affect Purkinje cells in the cerebellum and neurons of the brainstem and cervical spinal cord together with demyelination in the cerebellar white matter.
Myocardial lesions were found to be present in a large proportion of WNV cases. A mild to severe multifocal lympho-histiocytic myocarditis often extends and involves the epicardium and the endocardium. Non-inflammatory changes may also be present in the form of disseminated myocytolysis, multifocal haemorrhage and mineralization. Brain and heart lesions may have different or inverse severity in the same bird, e.g. cases with severe heart lesions may present mild or minimal brain involvement, and vice versa. This might reflect the acute or prolonged clinical course of the disease in affected individuals.
Lymphoid depletion is frequently found in the spleen, along with multifocal proliferation of reticulocytes and sometimes with signs of necrotizing splenitis. Some cases may exhibit multifocal interstitial lympho-histiocytic nephritis. Liver lesions range from multiple, focal haemorrhages and interstitial lympho-histiocytic inflammatory foci, sometimes containing heterophilic granulocytes, to acute multifocal coagulative necrosis of hepatocytes and moderate to severe congestion. Lungs are usually congested, with multifocal, acute haemorrhages and sometimes associated with a secondary bronchopneumonia. Mild lymphocytic enterocolitis and proventriculitis is a frequent feature accompanying WNV infection in birds.
The presence of WNV antigen in lesions can be demonstrated with immunohistochemistry (IHC) by using specific antibodies. The organs demonstrating the most pronounced immunolabelling are the brain and spleen, followed by the heart, kidneys, sciatic plexus and small intestine. In some cases viral antigen can also be visualized in other organs such as liver, lung, trachea, pancreas and thyroid gland. WNV antigen is most often observed in the cytoplasm of neurons, glial cells of the brain (Figure 9.2), macrophages, blood monocytes, myocardiocytes, fibro- cytes, tubular epithelial cells of the kidney, endothelial cells and smooth muscle cells of arteries, the epithelial cells lining the air capillaries of the lungs, enterocytes, pancreatic exocrine cells, hepatocytes, smooth muscle cells of the lamina muscularis mucosae in the small intestine and follicular epithelial cells of the thyroid gland.
Clinical signs and treatment
Birds are the principal hosts of WNV, and clinical manifestations of WNV infection range from subclinical infection, through a wide range of clinical signs to death. European wild bird species appear to be predominantly clinically unaffected by WNV infection. This may be attributed to their long co-adaptation to infection with the virus. However, as certain bird species do exhibit increased susceptibility to WNV infection it seems likely that spo-
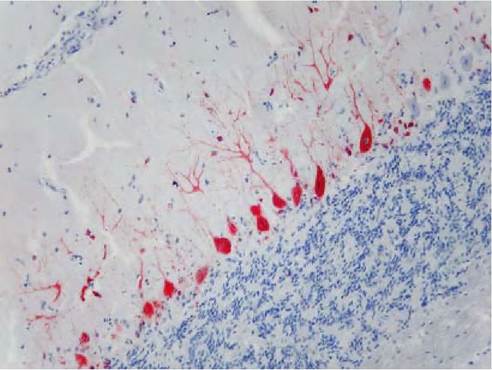
FIGURE 9.2 WNV antigen (red) in the cerebellum of a goshawk (positivity in neuronal axons and Purkinje cells). Immunohistochemistry (WNT 15R4 monoclonal antibodies, Horseradish-peroxidase labelled polymer (EnVision+ Kit), Mayer’s haematoxylin counterstain, 200?).
radic or small-scale disease events and mortality caused by WNV in Europe have not been recognized in the past.
Most bird infections are subclinical or produce mild signs of disease. However, these cases are also believed to result in high-level viraemias (as established for house sparrows and house finches ( Carpodacus mexicanus) in North America) and so serve as important reservoir and amplifying hosts ofWNV. Susceptible species, such as the goshawk and the sparrowhawk in Europe may be more seriously affected by the disease, initially showing depression, ataxia, anorexia, impaired vision and rapid loss of body condition, eventually developing more obvious neurological signs such as head tremors, paresis and seizures. The duration of clinical disease varies from an acute, fatal disease lasting for only one or two days, to a more prolonged illness that may lead to either death or recovery. Complete recovery may ensue in captive birds that are provided with supportive therapy and supplementary feeding as the neurological signs gradually subside.
In most mammals, including humans, clinical signs — if present — range from a mild, transient, flu-like, often febrile illness, to a neurological disease usually indicative of encephalomyelitis. The general symptoms may comprise headache, fatigue, body aches, swollen lymph nodes, fever and occasionally abdominal rash, and the neurological signs may include neck stiffness, stupor, disorientation, coma, tremors, convulsions, muscle weakness and paralysis.
In the absence of wildlife cases, horses can be significant indicators of local WNV activity. About 10% of WNV- infected horses show signs of clinical illness — comprising primarily generalized clinical signs, ataxia and sometimes more severe neurological signs — which is fatal in 20—40% of clinical cases.
Treatment protocols for clinical WNV cases are limited. Supportive therapy and supplementary feeding have been found to be essential to maximize the chances of recovery of individual patients from WNV infection. Captive, usually older, goshawks that survive the acute phase of the infection and maintain good body condition are found to recover from the infection, although chronic lesions resulting in impaired vision or minor neurological disorders may remain in some cases.
DIAGNOSIS
Direct demonstration of WNV in an infected host can be performed by virus isolation or RT-PCR. In the first case, the viable virus is isolated on cell cultures (e.g. Vero, RK13 or mosquito cells) either directly from host tissue samples or indirectly following mouse inoculation. The identification of the isolated virus may be performed by monoclonal antibodies (MAB) in virus neutralization, enzyme immunoassay, immunofluorescence assay or by RT-PCR.
RT-PCR can also be used to demonstrate directly the presence of viral RNA in clinical or pathological samples. The virus can be directly demonstrated in clinical blood samples (serum) during viraemia, cerebrospinal fluid, pharyngeal and cloacal swabs of birds or in fresh samples of the brain, spleen, heart and other parenchymal organs. Further identification and characterization of virus isolates may require the sequencing of amplified PCR products or full viral genomes.
Aggregates of WNV- labelled virions can be visualized in situ in infected organs by IHC performed on formalin- fixed tissue samples using WNV specific monoclonal or polyclonal antibodies. IHC may help elucidate the pathogenesis of WNV infection in different host species and their role in the epidemiology of WNV, also providing evidence for the pathogenicity of WNV strains by demonstrating and quantifying the association of the virus with lesions.
Neutralizing IgM and IgG antibodies to WNV can be detected by several serological methods. Considering the potential need for the testing of a wide range of bird and mammal species, the suitability of the test used for surveys or diagnostic purposes must be assessed beforehand. One such convenient test is a commercially available capture enzyme-linked immunosorbent assay (ELISA), which can be used on multiple species. Because of the high possibility of cross-reactions between flaviviruses in ELISA tests the diagnosis of WNV infection should be confirmed by the state-of-the-art plaque reduction neutralization assay (PRNT) performed in Vero cells that is specific for WNV. However, since a live virus strain is used in PRNT, it can only be performed in biosafety level 3 high- contingency laboratory facilities. Other tests that can be used for WNV serology are the haemagglutination inhibition test (HI), indirect immunofluorescence antibody test (IFAT) and microsphere immunoassay (MIA).
MANAGEMENT, CONTROL AND REGULATIONS
As the transmission cycle of WNV depends primarily on the presence and abundance of competent mosquito vectors, one of the recommended measures to reduce the risk of infection in urban and suburban areas is to control and limit mosquito breeding sites such as ditch water, small receptacles containing stagnant water and small water bodies, and to prevent mosquito bites by the use of repellents, netting and other methods. To the author’s knowledge, no attempts have been made so far to control WNV infections in wild animal populations. A passive surveillance scheme of human and horse encephalitis cases, complemented by passive surveillance of bird mortality and sentinel birds in endemic areas is recommended by the EU Scientific Committee on Veterinary Measures Relating to Public Health.
Extra-label vaccination — that is, the use of vaccine for species not covered by the manufacturer’s licensed instruction, with inactivated or recombinant WNV vaccines designed to be used in horses has been applied in captive individuals belonging to susceptible bird species (e.g. birds of prey). The efficacy of vaccination seems to vary on a wide scale, and it is apparently influenced by the type and make of the vaccine as well as the specific host species.
Animal cases of WNV infection are reportable to the World Organisation for Animal Health (OIE), whereas the human cases are reported within the EU and an epidemiological assessment of the WNV situation is performed by the ECDC.
PUBLIC HEALTH CONCERN
WNV is a zoonotic agent causing mainly sub-clinical infections in humans, but the infection develops into a flu-like illness (West Nile fever) in about 20% of human cases, and fatal encephalitis is recorded in about 1% of infected individuals. Immunocompromised and elderly people are at a higher risk of clinical infection. Severity of the disease may depend on the pathogenicity of the virus strain, and in this regard both the lineage 1 strain expanding through North America and the lineage 2 strain circulating in Central Europe and Greece are considered highly neuroinvasive.
SIGNIFICANCE AND IMPLICATIONS FOR ANIMAL HEALTH
Apart from humans, horses are the mammalian species most significantly affected by WNV infection. The severity of WNV-induced neurological disease may vary greatly and can lead to mortality. WNV is therefore considered an increasingly important disease of horses, and equine cases are also important indicators of WNV presence.
The current knowledge about the effect of WNV on European wild bird species is fairly limited. As clinical disease and mortality have so far only been recorded in rare cases and in a few species, the situation seems to be in stark contrast with the North American experience following the introduction of the virus. Focal impacts on the breeding success and population sizes of susceptible species such as the goshawk and sparrowhawk, mainly through increased fledgling mortality, may be expected in areas with high seasonal viral activity. The exposure of hitherto unexposed European bird populations through the potential expansion of the range and intensity of WNV circulation may result in unforeseen mortality events in the above-mentioned or other highly susceptible northern species such as the gyr falcon.