Changes in community structure
As we'll discuss in Chapter 16, ecological communities can be characterized by the number and relative abundances of the species they contain as well as by physical features of the environment.
Parasites can alter and be altered by each of these aspects of communities.In this chapter, we have seen several cases in which a parasite reduced the abundance, or even the geographic range, of its host, and we have also seen that parasites can change the outcome of species interactions. Such changes can have profound effects on the composition of communities. For example, a parasite that attacks a dominant competitor can suppress that species, causing the abundances of inferior competitors to increase. Such an effect was observed in six stream communities studied by Kohler and Wiley (1997). Prior to recurrent outbreaks of a fungal pathogen, the caddisfly Glossosoma nigrior was the dominant herbivore in each of the six communities. The fungus devastated Glossosoma populations, reducing their densities nearly 25-fold, from an average of 4,600 individuals per square meter to an average of 190 individuals per square meter. This drastic reduction in Glossosoma density allowed increases in the abundances of dozens of other species, including algae, grazing insects that ate algae, and filter feeders such as blackfly larvae. In addition, several species that previously were extremely rare or absent from the communities were able to establish thriving populations, thus increasing the diversity of the communities.
Parasites can also cause changes in the physical environment. This can happen when a parasite attacks an organism that is an ecosystem engineer, a species whose actions change the physical character of its environment, as when a beaver builds a dam (see Concept 16.3). As we learned earlier in Concept 13.4, the amphipod Corophium volutator can function as an ecosystem engineer in its tidal mudflat environment: in some circumstances, the burrows it builds hold the mud together, preventing the erosion of silt and causing the formation of “mud islands” that rise above the surface of the water at low tide.
As described earlier, trematode parasites can drive local Corophium populations to extinction (FIGURE 13.19A). When this happens, erosion rates increase, the silt content of the mudflats decreases, and the mud islands disappear (FIGURE 13.19B-D). Along with these physical changes, in one instance, the abundances of ten large species in the mudflat community changed considerably in the presence of the parasite, including one species (a ribbon worm) that was driven to local extinction (K. N. Mouritsen, personal communication).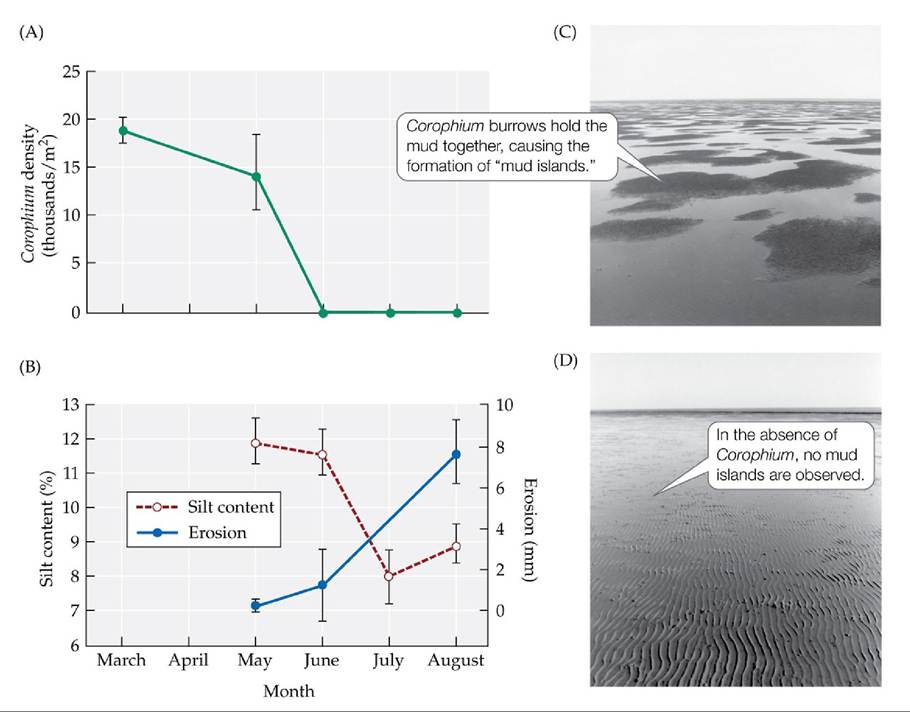
FIGURE 13.19 Parasites Can Alter the Physical Environment Infectionoftheamphipod Corophium volutator by a trematode parasite affects not only the host, but its entire tidal mudflat community. (A) The trematode can drive amphipod populations to local extinction. (B) In the absence of Corophium, the erosion rate increases and the silt content of the mudflats decreases. (C,D) The overall physical structure of the mudflats also changes [compare (C) with (D)]. Error bars show ± one standard error of the mean. (After K. N. Mouritsen et al. 1998. JMarBiolAssoc U.K. 78: 1167-1180; K. N. Mouritsen and R. Poulin. 2002. Parasitology 124: S101-S117.) View larger image
Finally, certain aspects of a community can be important in pathogen success and disease transmission. As we will learn in the Case Study in Chapter 19, the species diversity within a community can reduce the emergence and transmission of infectious diseases in wildlife and humans.
Climate Change Connection
Climate Change and Disease Spread
As we saw in Chapter 12, changes in climate are expected to have wide-ranging effects on species interactions and ultimately ecological communities (see Figure 12.22). For example, because mosquitoes and other vectors (organisms that transmit pathogens from one host to another) are often more active or produce more offspring under warm conditions, scientists have predicted that ongoing climate change may cause the incidence of some diseases to rise in human and wildlife populations (Epstein 2000; Harvell et al.
2002).A growing body of evidence supports this prediction. In one such study, increases in ocean temperatures were strongly correlated with increases in coral diseases along Australia’s Great Barrier Reef (Bruno et al. 2007). Similar results have been found in corals at other locations, as well as in a variety of amphibian and shellfish populations (Harvell et al. 2009).
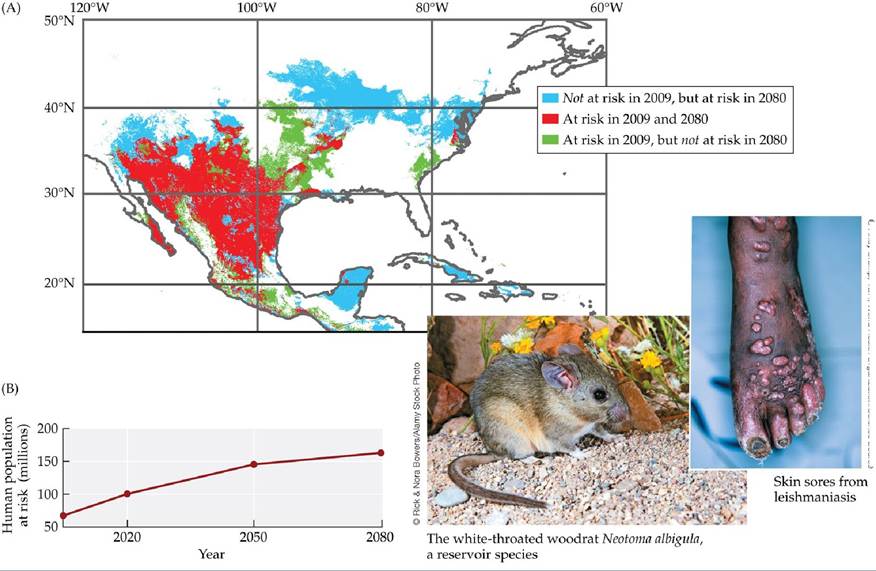
Climate change is also expected to change the distributions of some pathogens and their vectors by changing the locations where conditions are suitable for those organisms. For example, Gonzalez et al. (2010) found that climate change is likely to increase the risk of leishmaniasis in North America by increasing the geographic ranges of its reservoir species (rodents in the genus Neotoma that can harbor the pathogen) and its sand fly vectors (FIGURE 13.20). Similarly, the number of people at risk from malaria, cholera, and the plague may increase as global temperatures continue to warm (see citations in Ostfeld 2009).
© Andy Crump, TDR, World Health OrganizationZScience Source
FIGURE 13.20 Climate Change May Increase the Risk of Leishmaniasis in North America Leishmaniasis can cause severe skin sores, difficulty breathing, immune system impairment, and other complications that can lead to death. There are currently 1 million new cases each year. Leishmaniasis is caused by protists in the genus Leishmania and spread by sand flies (bloodsucking insects in the genera Lutzomyia and Phlebotomus). In addition to infecting humans, the pathogen can persist in several reservoir species (rodents in the genus Neotoma). (A) Change in the geographic regions in which people are predicted to be at risk from leishmaniasis due to the presence of at least one vector and reservoir species. (B) Change in numbers of people predicted to be at risk due to the presence of at least one vector and reservoir species. (After C.
Gonzalez et al. 2010. PLOS Neglected Trop Dis 4: 1-16.) View larger imageOverall, these studies and others (see Tylianakis et al. 2008; Gilman et al. 2010) indicate that ecological interactions will influence how future climate change will affect the incidence of disease in humans and many other species. Likewise, Costello et al. (2009) outlined major threats to human health from direct and indirect effects of climate change on disease incidence, food and water insecurity, and extreme climate events (such as hurricanes and floods that create
conditions that favor the spread of diseases). It is highly likely that climate change will also have direct and indirect effects on the incidence of disease in the populations of many species other than humans, contributing to the ongoing biodiversity crisis.
A Case Study Revisited
Enslaver Parasites
Returning to a question that we posed in the Case Study, how do enslaver parasites manipulate the behavior of their hosts? In some cases, we have hints of how they do this. Consider the tropical parasitoid wasp Hymenoepimecis argyraphaga and its host, the orbweaving spider Plesiometa argyra. The larval stage of this wasp attaches to the exterior of a spider's abdomen and sucks the spider's body fluids. When fully grown, the wasp larva induces the spider to make a special “cocoon web” (FIGURE 13.21). Once the spider has built the cocoon web, the larva kills and eats the spider. The larva then spins a cocoon and attaches it to the cocoon web. As the larva completes its development within the cocoon, the cocoon web serves as a strong support that protects the larva from being swept away by torrential rains.

FIGURE 13.21 ParasitesCanAlterHostBehavior Theparasitoidwasp
Hymenoepimecis argyraphaga dramatically alters the web-building behavior of the orbweaving spider Plesiometa argyra. (A) The web of an uninfected spider.
(B) The “cocoon web” of a parasitized spider. (After W. G. Eberhard. 2001. JArachnol 29: 354.) View larger imageA parasitized spider builds normal webs right up to the night when the wasp induces it to make a cocoon web. This sudden change in the spider's web-building behavior suggested that the wasp might inject the spider with a chemical that alters its behavior. To test this idea, William Eberhard (2001) removed wasp larvae from spider hosts several hours before the time when a cocoon web would usually be made. Wasp removal sometimes resulted in the construction of a web that was very similar to a cocoon web, but more often resulted in the construction of a web that was intermediate in form but differed substantially from both normal and cocoon webs. In the days that followed the removal of the parasite, some spiders partially recovered the ability to make normal webs. These results are consistent with the idea that the parasite induces construction of a cocoon web by injecting a fast-acting chemical into the spider. The chemical appears to act in a dose-dependent manner; otherwise, we would expect spiders exposed to the chemical to build only cocoon webs, not webs that are intermediate in form. Spiders build cocoon webs by repeating the early steps of their normal web-building sequence a large number of times; thus, the chemical appears to act by interrupting the spiders' usual sequence of web-building behaviors.
Other enslaver parasites also appear to manipulate host body chemistry. In the Case Study, we described hairworm parasites that cause crickets to commit suicide by jumping into water. Thomas and colleagues (2003) have shown that the hairworm causes biochemical and structural changes in the brain of its cricket host. The concentrations of three amino acids (taurine, valine, and tyrosine) in the brains of parasitized crickets differ from those in crickets that have not been parasitized. Taurine, in particular, is an important neurotransmitter in insects, and it also regulates the brain's ability to sense a lack of water.
Hence, it is possible that the parasite induces its host to commit suicide by causing biochemical changes in its brain that alter the host's perception of thirst.The papers by Eberhard (2001) and Thomas et al. (2003) suggest that some parasites enslave their hosts by manipulating them chemically. But even in Eberhard's work, which indicates that the wasp injects a chemical into its spider host, the chemical in question has not been found. If this chemical were known, it could be injected into unparasitized spiders; if those spiders constructed cocoon webs, we would have a clear understanding of how the parasite manipulates the spider.
Although a definitive chemical experiment such as this has yet to be performed, a similar genetic experiment was performed for spongy moths (Lymantria dispar) enslaved by a virus (Hoover et al.
2011). Spongy moths infected with this virus move to the tops of trees shortly before they die; after death, the bodies of the moths liquefy and release millions of infective viral particles. Uninfected spongy moths do not exhibit this climbing behavior before death. Based on previous work, Hoover and colleagues hypothesized that the expression of a particular viral gene (the egt gene) caused infected moths to move to treetops shortly before death. In a laboratory test of this hypothesis, they found that moths infected with the typical, or wild-type, virus strain died at higher positions than did moths infected by viruses from which the suspect gene had been removed (FIGURE 13.22)—strong evidence that they had succeeded in identifying the first known “enslaver gene.”
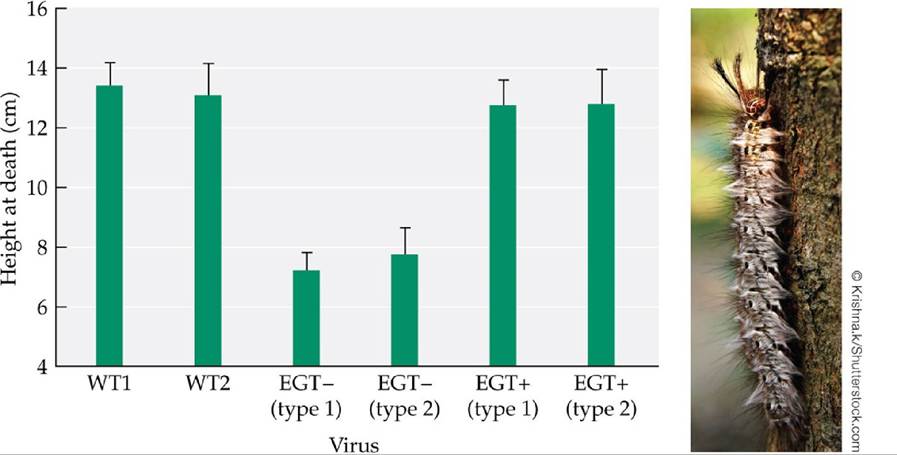
FIGURE 13.22 A Parasite Gene That Enslaves Its Host Spongy moths infected by a virus (Lymantria dispar nucleopolyhedrovirus, or LdMNPV) climb to high locations before they die—a behavior that benefits the virus but not the moth. To test the hypothesis that a particular viral gene (the egt gene) affects this behavior, researchers reported the height at death of spongy moth caterpillars reared in cages and subjected to the following treatments: WT viruses (two different natural, or wild-type, viruses); EGT- viruses (two different experimental viruses from which the egt gene had been removed); and EGT+ viruses (two different experimental viruses from which the egt gene was first removed, then
replaced). Error bars show one standard error of the mean.
Explain why the researchers included the WT and EGT+ treatments.
(After K. Hoover et al. 2011. Science 333: 1401.) View larger image
Connections in Nature
From Chemicals to Evolution and Ecosystems
Enslaver parasites that manipulate their hosts exert strong selection pressure on host populations, so resistance to the manipulations of enslaver parasites might be expected to evolve in host populations. For example, selection would favor host individuals with the ability to recognize and destroy chemicals that a parasite uses to alter host behavior. Likewise, enslaver parasites might be expected to evolve the ability to overcome host resistance mechanisms.
To date, we know of no such evidence of ongoing host-enslaver parasite coevolution. However, interactions between enslaver parasites and their hosts do provide evidence of previous evolutionary change. Like any parasite, an enslaver parasite has adaptations that allow it to cope with host defenses (otherwise it would not survive). More specifically, an enslaver parasite that uses a chemical to manipulate a specific host behavior is beautifully adapted to take advantage of the body chemistry of its host. Such evolutionary links between enslaver parasites and their hosts illustrate a central feature of both ecology and evolution: ecological interactions affect evolution, and vice versa, at times making it difficult to distinguish one from the other (see Concept 6.5). As we've seen in this chapter, the outcome of such ecological and evolutionary interactions can have profound effects on individuals, populations, communities, and ecosystems. As evolutionary change tips the balance back and forth, first in favor of the host, then in favor of the parasite, we can expect concomitant changes in the population dynamics of other species, such as those that compete with or eat the host or the parasite. Viewed in this way, communities and ecosystems are highly dynamic, always shifting in response to the ongoing ecological and evolutionary changes that are occurring within them.