Early studies suggested that resource partitioning was the main mechanism of coexistence
As we learned earlier from the two-species studies of Gause (1934a) on Paramecium (see Concept 14.2) and Connell (1961a,b) on barnacles (see Concept 14.4), species that compete with each other may coexist by using slightly different resources.
Robert MacArthur, whose work on the equilibrium theory of island biogeography we described in Concept 18.3, played a pioneering role in understanding how this principle might be applied to whole communities, where multiple species interactions are occurring all at once.MacArthur studied warblers, small and brightly colored birds that co-occur in the forests of northern North America. The idyllic New England forests that MacArthur studied are home to an array of warbler species, Setophaga spp. (renamed from Dendroica spp.) that migrate from the tropics each spring to breed and feed on insects. Through a series of detailed natural history observations in the summers of 1956 and 1957 in Maine and Vermont, MacArthur (1958) recorded the feeding habits, nesting locations, and breeding territories of five species of warblers to find out how they might coexist in the face of very similar resource needs.
MacArthur began mapping the locations of warbler activity in tree canopies and found that the warblers were using different parts of the habitat in different ways (FIGURE 19.9). For example, yellow-rumped (S. coronata) warblers fed from the middle parts of trees to the forest floor, while bay-breasted (S. castanea) and black-throated green (S. virens) warblers fed more in the middle of a tree, both inside and toward the outside of the tree canopy. Blackburnian (S. fusca) and Cape May (S. tigrina) warblers both fed on the outside tops of trees, often catching their prey in midflight. MacArthur found that the nesting heights of the five warbler species also varied, as did their use of breeding territories.
Taken together, these observations supported his hypothesis that the warblers, although using the same habitat and food resources, were able to coexist by partitioning those resources in slightly different ways. MacArthur's work, which was part of his PhD thesis, earned him the prestigious Mercer Award, bestowed each year for the best paper in ecology.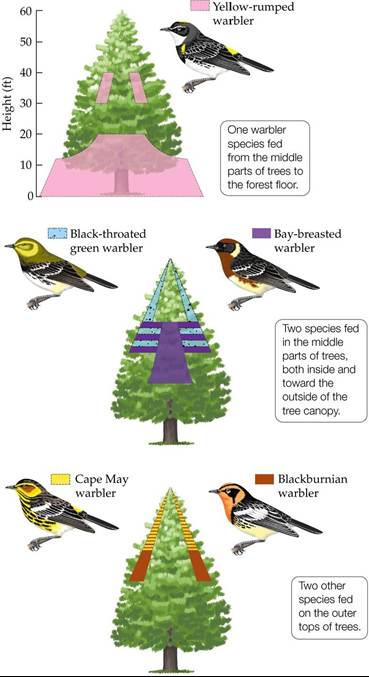
FIGURE 19.9 Resource Partitioning by Warblers Robert MacArthur studied the habitat and food choices of five species of warblers in New England forests. He found that the warblers partition resources by feeding in different parts of the same trees. The colored shaded areas in each tree diagram represent the parts of trees where each warbler species fed most often. (After R. H. MacArthur. 1958. Ecology 39: 599-619.) View larger image
MacArthur, along with his brother John MacArthur (MacArthur and MacArthur 1961), extended these ideas about resource partitioning in a study of the relationship between bird species diversity (calculated using the Shannon index; Concept 16.2) and foliage height diversity (a measure of the number of vegetation layers in a community that serves as an indication of habitat complexity, also calculated using the Shannon index). They found a positive relationship between the two in 13 tropical and temperate bird habitats from Panama to Maine (FIGURE 19.10). Interestingly, bird species diversity was not related to plant diversity per se, beyond the effects of foliage height diversity, suggesting that tree species identity was less important than the structural complexity of the habitat.
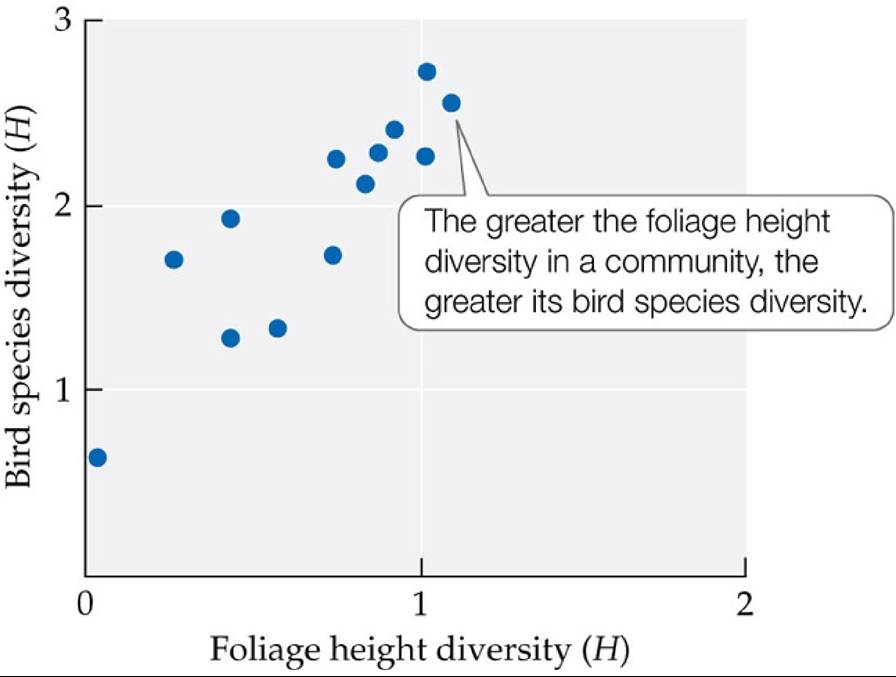
FIGURE 19.10 Bird Species Diversity Is Higher in More Complex Habitats MacArthur and MacArthur plotted bird species diversity against foliage height diversity (a measure of habitat complexity) for 13 different communities. Both kinds of diversity were calculated for each community, using the Shannon index (H).
(After R. H. MacArthur and J. W. MacArthur. 1961. Ecology 42: 594-598.) View larger imageAnother important resource partitioning study comes from phytoplankton communities. In Concept 14.1, we learned about David Tilman and colleagues' (1981) study of two species of diatoms that competed for silica (which diatoms use to build their cell walls). When the two species were grown together in a laboratory environment with limited supplies of silica, one outcompeted and excluded the other (see Figure 14.6). How, then, do diatom species coexist in nature? Tilman (1977) proposed what has become known as the resource ratio hypothesis, which posits that species coexist by using resources in different ratios or proportions. He predicted that diatoms would be able to coexist, despite using the same set of limiting nutrients, by acquiring those nutrients in different ratios. By growing two diatom species, Cyclotella and Asterionella, in laboratory environments that differed in their ratios of silica (SiO2) to phosphorus (PO4), Tilman found that Cyclotella was able to dominate only when the ratio of silica to phosphorus was low (approximately 1:1). When the ratio of silica to phosphorus was high (more like 1,000:1), Asterionella outcompeted Cyclotella. Only when the ratios of silica and phosphorus were limiting to both species (in the range of 100:1 to 10:1) could they coexist. Even though both species needed the same set of nutrients, it was the way in which they partitioned those resources that allowed them to coexist.
Outside of a laboratory setting, this type of partitioning would work best if resources naturally varied within the environment. What is the support for this possibility in the field? In a detailed survey, Robertson and colleagues (1988) mapped resource distribution in an abandoned agricultural field in Michigan that had been colonized by grassland plants. They found considerable variation in soil nitrogen and moisture at spatial scales of a meter or less (FIGURE 19.11).
These patches of nitrogen and water resources did not necessarily correspond to topographic differences, and they were not correlated with each other. If we were to overlay the nitrogen map on the water map, we would find even smaller patches corresponding to different proportions of these two resources. Some of the best evidence of resource partitioning in plants comes from experiments that manipulate species richness and measure productivity, as we will explore in more detail in Concept 19.4.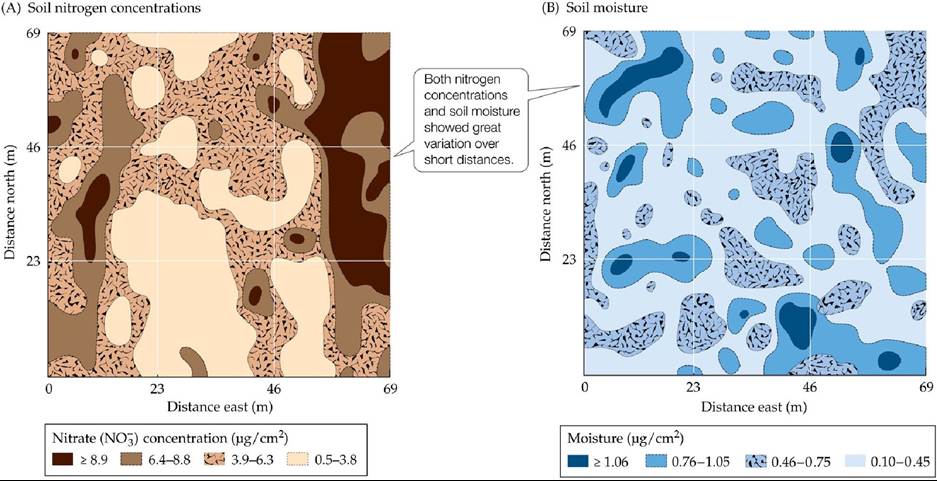
FIGURE 19.11 ResourceDistributionMaps Mapping of (A) nitrogen concentrations and (B) soil moisture in an abandoned agricultural field revealed considerable small-scale variation (μg∕cm2 = micrograms per square centimeter). (From G. P. Robertson et al. 1988. Ecology 69: 15171524.) View larger image
The theory of resource partitioning relies on the assumption that species have evolved mechanisms for using resources in different, but complementary, ways, thus increasing their ability to coexist. As we learned in our discussion of species interactions in Unit 4, there are numerous other processes that can alter the outcome of species interactions and allow coexistence. In the next section, we will consider how those processes control species diversity at the local scale.