Other types of environmental variation
Variation in the consumer mortality rates, which is included in eqs (9.1) and (9.2), can also affect coexistence. Such variation could occur in conjunction with nonlinear functional and/or numerical responses.
In the linear consumer models treated in Chapter 8, cycles in consumer mortality had no effect on the mean resource abundance, and thus, no effect on coexistence. This is no longer the case when one or both of the consumers’ per capita growth rates are nonlinear and this holds for both numerical and functional responses. There are many possibilities for such variation; it can occur in one or both species, and in the latter case it may differ in phase or amplitude between species. If the resource growth rate also varies, there are even more possible combinations of different types of variation. This section will present a few examples to illustrate some of circumstances that have the most obvious effects on coexistence. It will only consider identical sinusoidal variation in one or both consumer species.9.5.1 Nonlinear numerical responses
I begin by considering correlated mortality in both consumers in the numerical response model, eqs (9.2); both mortality rates vary between zero and twice their mean. The fact that the baseline consumer per capita mortality rates are low relative to the baseline resource per capita growth rate in the system considered here means that a longer period q is required to generate enough variation in resource abundance to have a significant effect on coexistence. Here, coexistence does not occur until q is considerably larger than 60, and the bandwidth is relatively narrow even for q = 100. When q = 60, not only is coexistence impossible for all d2, but there are alternative exclusion outcomes for 0.335 < d2 < 0.426. When q = 100, coexistence is the only outcome for the relatively narrow range of 0.351 < d2 < 0.424.
A small additional range of mortalities (0.424 < d2 < 0.449) allows alternative outcomes of coexistence or exclusion of consumer 2 by consumer 1. Even if we include this additional range, the total coexistence bandwidth is only about 1/3 that of the comparable model with resource variation (eqs 9.1) in which species 1 has type IIfunctionalresponse (Table 9.1). Ifthe period q is raised to 150, this yields coexistence for 0.313 < d2 < 0.450, with exclusion of species 2 above this range, and exclusion of 1 below it; no alternative attractors were observed. If the linear consumer species 2 is the only one that exhibits variable mortality, coexistence is not possible. There are alternative exclusion outcomes depending on initial abundances when 0.150 < d2 < 0.313. Mortalities d2 lower than this range only allow species 2 to persist, while higher d2 values only allow species 1 to persist. If only the nonlinear species 1 exhibits mortality variation, coexistence occurs for 0.150 < d2 < 0.451, with exclusion of species 2 above this range, and exclusion of 1 below it.The above comparison of single and dual consumer mortality variation can be understood qualitatively based on the general requirement that, in order for two species to coexist, at least one species must have some stabilizing process that decreases its relative competitive ability as its short-term mean abundance increases. If variable mortality is associated with the nonlinear consumer 1 in eqs (9.2), this will lead to greater resource fluctuations as its numbers increase, which will slow its growth and provide an opportunity for the linear species to coexist. Thus, variation in the mortality of consumer 1 produces a relatively wide coexistence bandwidth. This is reduced, but not eliminated by having mortality variation in the linear consumer as well. A numerically dominant linear species 2 produces large amplitude fluctuations in the resource that often give it the ability to exclude species 1.
9.5.2 Nonlinear functional responses
The comparable model with species 1 having a nonlinear functional response (Table 9.1 above) also yields somewhat different results when consumer mortality variation is substituted for resource growth variation. In the absence of any environmentally caused cycles, consumer 1’s type II response implies a coexistence bandwidth for the linear species of d2 = 0.15-0.507. Coexistence was examined in this variable consumer mortality model for periods of 15 and 30 to provide a comparison with the case of variation in resource growth rates in Table 9.1. Somewhat less detailed results for q = 5 and q = 100 are given at the end of this section.
For q = 15, coexistence was always observed from d2 = 0.286 through d2 = 0.516. This compares to the much narrower bandwidth of 0.502 < d2 < 0.539 for the model considered earlier that only has variation in resource growth. Note that this coexistence attractor in the resource-variation model occurred in conjunction with an alternative attractor characterized by exclusion of consumer 2. Thus, variation driven by fluctuations in both consumers’ mortalities was considerably more favourable for coexistence. However, this is largely because the smaller amplitude of consumer mortality cycles compared to resource growth cycles results in less interference of the environmentally driven cycles on the interaction-driven cycles.
If q = 30 for the consumer mortality variation, coexistence is possible from d2 = 0.450 to d2 = 0.535. Exclusion of consumer 1 always occurred below this range,
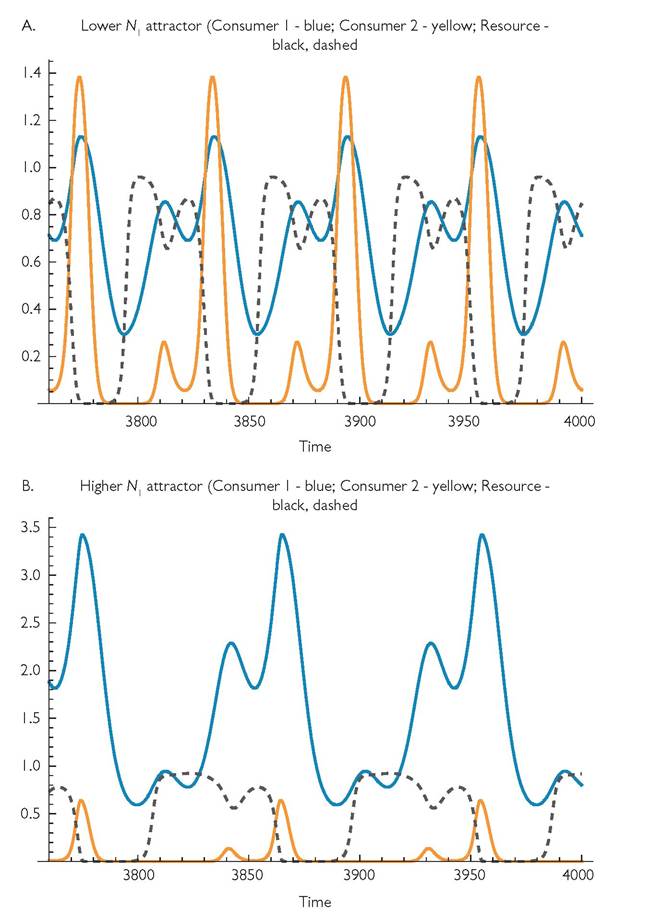
Fig. 9.5 An example of alternative coexistence outcomes that occur with the nonlinear functional response model having variation in both consumer mortality rates. The parameters are: r = 1; k = 1; Ci = 1, C2 = 1; hl = 10, h2 = 0; B1 = 1, B2 = 1; di = 0.06, d2 = 0.5; m = 0.0001; q = 30, γp1 = 1, γp2 = 1.
Note the difference in the scale of the ó-axis, due to the much greater maximum abundance of consumer 1.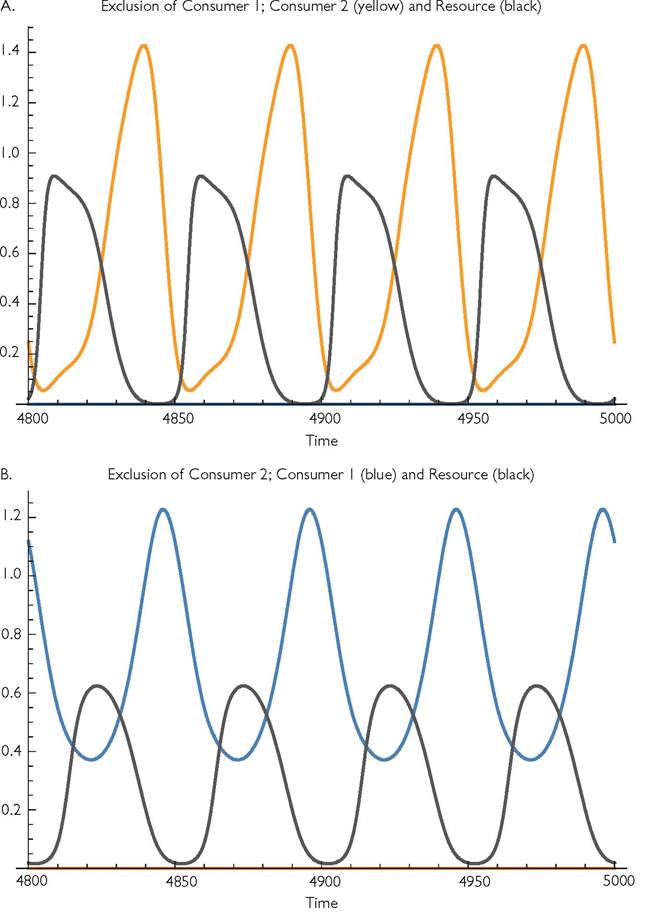
Fig. 9.6 'The alternative attractors for eqs (9.2) with variation in both consumers’ mortality rates. The parameters are: r = 1; k = 1; Ci = 1, C2 = 1; h1 = 10, h2 = 0; Bi = 1, B2 = 1; d1 = 0.06, d2 = 0.4; m = 0.0001; q = 50, γp1 = 1, γp2 = 1. Alternative exclusion outcomes exist for values of d2 between 0.29625 and 0.545.
and exclusion of consumer 2 occurred above this range. This compares to the much wider range of mortalities allowing coexistence (0.340-0.697; see Table 9.1) in the comparable case of variation in resource growth rate with q = 30. This is again related to the fact that the environmentally caused variation is of greater amplitude in the case of resource growth variation than in the case of consumer mortality variation. Unlike the preceding example (q = 15), q = 30 has a very positive effect on coexistence. The bandwidth noted above actually overstates the possibility of coexistence in the consumer mortality variation model because one or more alternative attractors are found for a wide range of different mortalities in the q = 30 case. Most significant from the standpoint of coexistence is the fact that exclusion of species 2 by species 1 is a possible outcome for values of d2 from 0.491 to the maximum of 0.536. Exclusion of species 1 by species 2 occurs as an alternative to coexistence for d2 values from 0.450 to 0.482. Thus, there is only a narrow range of mortalities of the linear species over which exclusion never occurs for any initial conditions (d2 from 0.482 to 0.491). In addition, there are parameter ranges over which two different coexistence attractors exist, and parts of these ranges also allow a third (exclusion) attractor.
Figure 9.5 illustrates one pair of alternative coexistence attractors that occur in the case of q = 30. Alternatives are usually characterized by different periods for the variation in abundances; these are different small integer multiples of the basic period. The period for Figure 9.5A is 60, while that of 9.5B is 90.Very short periods (e.g. q = 5) in this consumer mortality variation model have relatively modest effects on coexistence bandwidth; the cycle driven by consumer 1 dominates the dynamics except when d2 is low enough to greatly reduce the abundance of consumer 1. Moderately long periods are particularly likely to have alternative exclusion outcomes. Figure 9.6 shows the two exclusion outcomes that are possible if the parameter set described in the previous paragraph has q = 50 and d2 = 0.4. This case allows alternative exclusion for a wide range of mortalities between d2 = 0.296 and d2 = 0.545. The alternatives illustrated in Figure 9.6 are characterized by quite different mean resource abundances. Still longer periods (e.g. q = 100) have narrower coexistence bandwidths than in the otherwise comparable model with resource-growth driven rather than consumer mortality driven cycles. For the system considered in the previous paragraph coexistence occurs for d2 between 0.36 and 0.59, which is roughly 1/3 the range for q = 100 under resource growth variation (see Table 9.1). This lower bandwidth is largely a consequence of the lower amplitudes of the resource fluctuations caused by consumer mortality rate variation compared to those caused by resource growth variation.
9.5.3 Remaining unknowns
This section definitely does not constitute an adequate analysis of the possible effects on competition between two consumers for a single resource with periodic variation in consumer mortality rates. Many options regarding different amplitudes or phase shifts between consumers, or combinations of consumer- and resource-driven cycles remain to be explored.
Obviously, different functional forms or even different parameter values should be examined. However, the examples provided serve to show that the dynamics do not fit well into a framework based purely on invasion, due to the frequent occurrence of alternative attractors. The results also are sufficient to show that differences in linearity do not necessarily promote coexistence in variable environments. Finally, they demonstrate that a considerable diversity of effects on coexistence can arise from relatively small changes in parameter values and from different sources of variation.Future work should examine: (i) various models of abiotic growth; (ii) biotic resource systems in which the resource’s own resource(s) are represented; (iii) models of combined abiotic and biotic growth; (iv) models with two or more potentially interacting resource species/types; and (v) models in which all species exhibit correlated seasonal variation in several parameters, as in most higher-latitude environments. The consequence of different dynamic rates of consumer and resource also needs more study. A wide variety of different functional response forms arise from adaptive behaviour (Chapter 3), and some of these are likely to lead to different impacts on coexistence in variable environments.
9.6