Positive interactions influence the abundances and distributions of populations
As examples earlier in this chapter suggest, mutualism and commensalism can provide benefits that increase the growth, reproduction, or survival of individuals in one or both of the interacting species—a point that was demonstrated recently for a defensive bacterial symbiont that increased the reproductive success of its fruit fly host (FIGURE 15.16).
As a result, mutualism and commensalism can affect the abundances and distributions of the interacting species. To explore these issues further, we will first examine how an ant-plant mutualism affects the abundances of its members. We will then consider how mutualism and commensalism influence the distributions of organisms.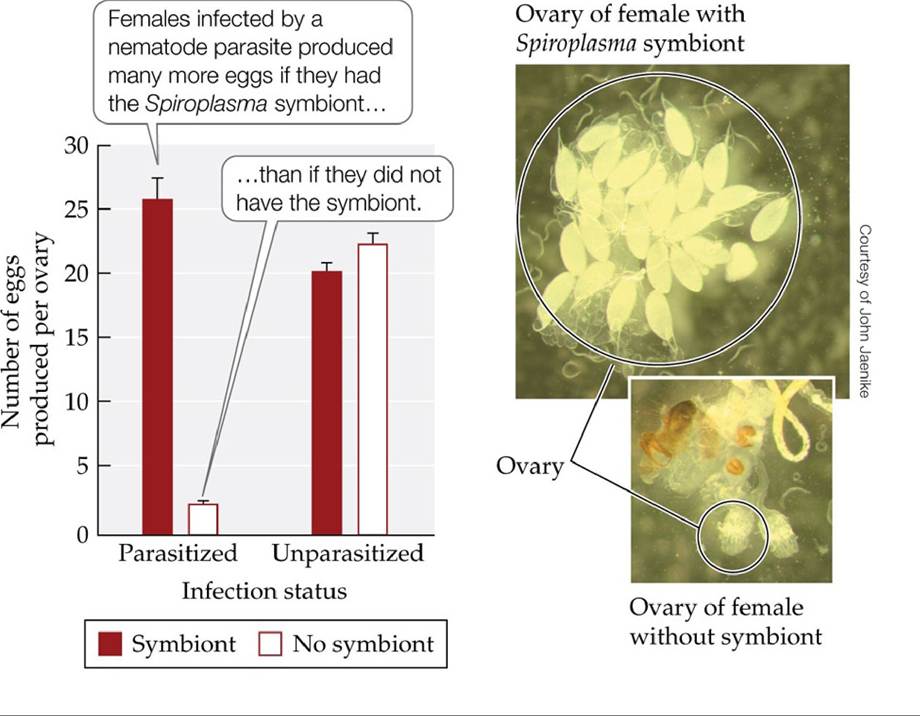
FIGURE 15.16 A Symbiont Increases the Fertility of Its Host Bacteriainthegenus
Spiroplasma are obligate symbionts that live within the cells of their host, the fruit fly Drosophila neotestacea. The graph shows the number of eggs produced by laboratory-reared female flies that either had Spiroplasma symbionts (red bars) or did not have Spiroplasma symbionts (white bars), and that either were infected by the nematode parasite Howardula (parasitized) or were not infected by it (unparasitized). Howardula can sterilize female flies and reduce the mating success of male flies. Error bars show one standard error of the mean. (After J. Jaenike et al. 2010. Science 329: 212-215.) View larger image
Effects on Abundance
The effects of mutualism on abundance can be seen in the mutualistic relationship between ants in the genus Pseudomyrmex spp. and the bullhorn acacia (Acacia spp.). This plant has unusually large thorns, which provide a home for the ants (FIGURE 15.17A). The thorns have a tough, woody covering but a soft, pithy interior that is easy for the ants to excavate.
A queen ant establishes a new colony by burrowing into a green thorn, removing some of its pithy interior, and laying eggs inside the thorn. As the colony grows, it eventually occupies all of the acacia's thorns.
© Niclxjlas SmytheZScjence Source
FIGURE 15.17 An Ant-Plant Mutualism (A) Acacia ants (Pseudomyrmex spinicola) tending to larvae and pupae inside an acacia thorn. (B) A nectary at the base of a leaf and Beltian bodies at the leaflet tips. (C) Ants have removed the plants that grew near this acacia, creating a competitor-free zone for the plant. View larger image
The ants feed on nectar, which the plant secretes from specialized nectaries, and on modified leaflet tips called Beltian bodies, which are high in protein and fat (FIGURE 15.17B). The ants aggressively attack insect and even mammalian herbivores (such as deer) that attempt to eat the plant. The ants also use their mandibles to maul other plants that venture within 10-150 cm of their home acacia, thus providing the acacia with a competitor-free zone in which to grow
(FIGURE 15.17C).
Do the services provided by the ants benefit the acacias? To find out, Dan Janzen removed ants from some acacia plants and compared the growth and survival of those plants with those of plants that had ant colonies. The results were striking. On average, bullhorn acacias with ant colonies weighed over 14 times as much as acacias that lacked colonies; acacias with ants also had higher survival rates (72% vs. 43%) and were attacked by insect herbivores much less frequently (Janzen 1966).
If a bullhorn acacia lacks an ant colony, the repeated loss of its leaves and growing tips to herbivores often kills the plant in 6-12 months. The ants, in turn, depend on the acacias for food and a home, and they cannot survive without these plants. Thus, the ant-acacia mutualism has considerable effects on the abundance of each partner.
Furthermore, both the ant and the plant have evolved unusual characteristics that benefit their partners. For example, Pseudomyrmex ants that depend on acacias are highly aggressive, remain active for 24 hours a day (patrolling the plant surface), and attack vegetation that grows near their home plants; Pseudomyrmex species that do not form mutualisms with acacias show none of these traits. Similarly, acacias that form mutualisms with ants have enlarged thorns, specialized nectaries, and Beltian bodies on their leaves; few nonmutualistic acacia species show these traits. Overall, both the ants and the acacias appear to have evolved in response to their partners, making the antacacia partnership an example of an obligate and coevolved mutualism.Effects on Distribution
There are literally millions of positive interactions in which one species provides another with favorable habitat and thus influences its distribution. Specific examples include corals that provide their algal symbionts with a home and fungal symbionts that enable plants to live in environments they otherwise could not tolerate (such as the example of the Curvularia fungi that enable the grass Dichanthelium to live in high-temperature soils). Of course, obligate mutualisms, such as the fig-fig wasp mutualism discussed earlier, have a profound influence on the geographic distribution of the interacting species because neither can live where its partner is absent.
It is very common for a group of dominant species, such as the trees in a forest, to determine the distributions of other species by physically providing the habitat on which they depend. Many plant and animal species are found only in forests. Such “forest specialists” either cannot tolerate the physical conditions of more open areas (such as a nearby meadow) or are prevented from living in those open areas by competition with other species. Similarly, at low tide in marine intertidal communities, many species (e.g., crabs, snails, sea stars, sea urchins, barnacles) can be found under the strands of seaweeds that are attached to the rocks. The seaweeds provide a moist and relatively cool environment that enables some species to live in higher regions of the intertidal zone than they otherwise could. Finally, many sandy and cobblestone beaches are stabilized by grasses such as Ammophila breviligulata and Spartina alterniflora. By holding the substrate together, these species enable the formation of entire communities of plants and animals.
Many forest specialists have little direct effect on the trees under which they live; hence, they have a commensalism with the trees of the forest. The same is true of many marine species that seek shelter under seaweeds and of many of the organisms that depend on substrate stabilization by grasses. In each of these cases, a positive interaction (often a commensalism) allows one species to have a larger distribution than it otherwise would.