Positive interactions can alter communities and ecosystems
The effects that commensalism and mutualism have on the abundances and distributions of species can affect other species interactions, which in turn can have consequences for community.
For example, if a dominant competitor depends on a facilitator, loss of the facilitator may reduce the performance of that dominant species and increase the performance of other species—thus changing the mix of species in the community or their relative abundances. As we'll see, when the structure of a community changes, properties of the ecosystem may also change.Community Diversity
Coral reefs are known for their astonishing beauty, and they are exceptional ecologically in that their fish communities are the most diverse vertebrate communities in the world. One of the most common interactions among these diverse coral reef fish is a service mutualism in which a small species (the “cleaner”) removes parasites from a larger fish (the “client”). The cleaner often ventures into the mouth of the client (FIGURE 15.18A). What prevents the client from simply eating the cleaner?
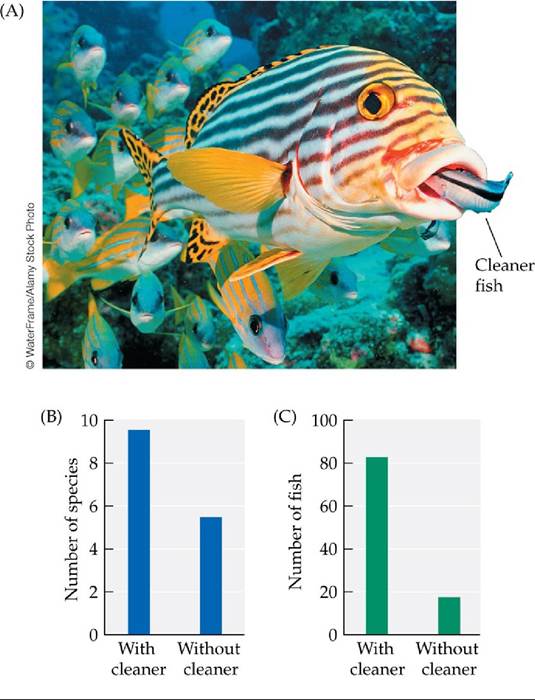
FIGURE 15.18 Ecological Effects of the Cleaner Fish Labroides dimidiatus (A) Looking for parasites, a cleaner fish places its head within the mouth of a much larger client, this sweetlips fish. The experimental removal of L. dimidiatus from small reefs within the Great Barrier Reef of Australia led to (B) a drop in the number of fish species found on the reefs and (C) a decrease in the total abundance of fish on the reefs. (B,C after A. S. Grutter et al. 2003. CurrBiol 13: 64-67.) View larger image
The answer appears to be that the benefit a client receives from cleaning (parasite removal) is greater than the energy benefit it could gain by eating the cleaner. In the Great Barrier Reef of Australia, individuals of the cleaner species Labroides dimidiatus were visited by an average of 2,297 clients each day, from which the cleaner removed (and ate) an average of 1,218 parasites per day (0.53 parasites per client).
To determine whether the activities of cleaners were translated into a reduction in the number of parasites found on clients, Alexandra Grutter experimentally removed L. dimidiatus from three of five small reefs.After 12 days, on the reefs from which the cleaners had been removed, there were 3.8 times more parasites on Hemigymnus melapterus fish than on the control reefs. In follow-up studies, Grutter and colleagues (2003) examined the effect of L. dimidiatus on the number of species and the total abundance of fish found on coral reefs. The results were dramatic: removal and exclusion of L. dimidiatus for a period of 18 months caused large drops in both the number of fish species and the total abundance of fish found on the reefs (FIGURE 15.18B,C).
Grutter's work shows that a mutualism can have a major effect on the diversity of species found in a community. Most of the species lost from the reefs without cleaners were species that typically move among reefs, including some large predators. Large predators can themselves affect the diversity and abundance of species, so the removal of cleaner fish could also result in further, but difficult- to-predict, long-term changes to the community.
Climate Change Connection
Climate Warming and Extinction of Plant-Pollinator Networks
As we have learned, plant-pollinator mutualisms are often highly specialized and have important benefits for both species involved in the interaction. These codependent species are also embedded within whole communities of plants and pollinators, connected through multiple mutualistic interactions. But what happens when climate change causes the extinction of one or more plant species within these networks, and what effects could that have on the extinction of pollinator species dependent on those plants? To explore this question, Jordi Bascompte, Maria Garcia, and
colleagues (2019) used species distribution models (see Concept
9.2 for use of species distribution models to predict the locations of chameleon species in Madagascar) to predict the extinctions and subsequent coextinctions of species due to warming temperatures within seven plant-pollinator networks across Europe, due to warming temperatures at two time horizons of 2050 and 2080 (FIGURE 15.19A).
The researchers were interested in measuring the response of plant-pollinator networks under two time horizons (2050 and 2080) and for two sources of extinction: (1) the direct effects of climate warming on plant species extinction rates and (2) the subsequent coextinction of pollinator species that depend on a mutualistic interaction with those plants. As FIGURE 15.19B reveals, there was a large latitudinal range in the rate of species extinctions, with the two Mediterranean networks showing higher numbers of species lost compared to the other northern networks for both 2050 and 2080. The differences were even greater when considering the additional coextinctions that were predicted to occur from the direct loss of plants to their pollinators (see Figure 15.19B). The results suggest that the Mediterranean plant-pollinator networks may be more vulnerable to climate change than the Eurosiberian networks. If so, the greater species loss in the Mediterranean networks could be the consequence of greater warming temperatures in southern regions but also a narrower distributional range of plants and pollinators in these networks, making it harder to “escape” to higher latitudes. This study suggests that it is important to consider the interdependencies among species when predicting extinction risk in the face of climate warming.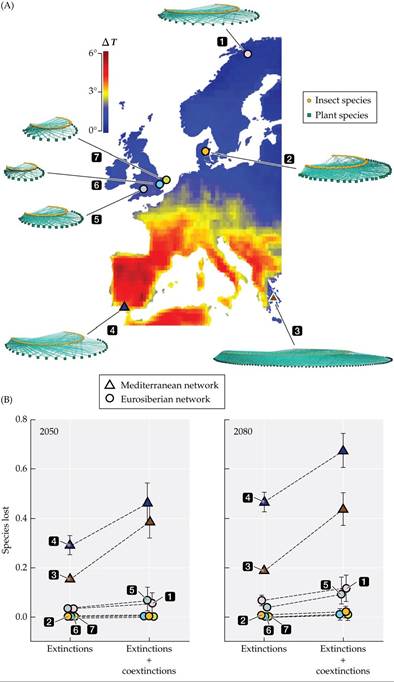
FIGURE 15.19 Plant-Pollinator Extinctions Predicted Under Climate Warming (A) Researchers measured the effects of the predicted increase in temperature (ΔT is the change in temperature in the warmest month from 2020 to 2080) on seven plant-pollinator networks (the green squares represent plants, the orange circles represent pollinator insects, and the solid lines represent interactions) across Europe. (B) Plant species extinctions and subsequent coextinctions of pollinators within each pollination network at two times in the future, 2050 and 2080.
Error bars show one standard error of the mean. The different numbers identify different networks, with the triangles representing the Mediterranean networks and the circles representing the Eurosiberian networks. (After J. Bascompte et al. 2019. SciAdv 5: eaav2539.) View larger imageSpecies Interactions and Ecosystem Properties
Barbara Hetrick and colleagues (1989) performed greenhouse experiments in which the presence of mycorrhizal fungi altered the outcome of competition between two prairie grasses, big bluestem (Andropogon gerardii) and junegrass (Koeleria macrantha). They found that big bluestem dominated when mycorrhizal fungi were present and that junegrass dominated when they were not. In a natural prairie community of which big bluestem was a dominant member, when David Hartnett and Gail Wilson (1999) suppressed mycorrhizal fungi with a fungicide, the performance of big bluestem decreased. At the same time, the performance of a variety of other plant species, including both grasses and wildflowers, increased. Hartnett and Wilson suggested that big bluestem’s dominance may have come from a competitive advantage conferred by its association with mycorrhizal fungi and that removal of those fungi removed that advantage and released the inferior competitors from the negative effects of competition.
Mycorrhizal associations can affect other features of ecosystems in addition to diversity, as shown in a 1998 study by Marcel van der Heijden, John Klironomos, and colleagues. In a large-scale field experiment, these scientists manipulated the number of species of mycorrhizal fungi (from 0 to 14 species) found in soils in which identical mixtures of the seeds of 15 plant species had been sown. After one growing season, plant dry weights and phosphorus content were measured. Plant root and shoot biomass increased as the number of species of fungi increased (FIGURE 15.20A,B), as did the efficiency of phosphorus uptake by plants (FIGURE 15.20C). These results show that mycorrhizal fungal species richness can influence key features of ecosystems such as net primary production (measured as the amount of new plant growth over one growing season) and the supply and cycling of nutrients such as phosphorus.
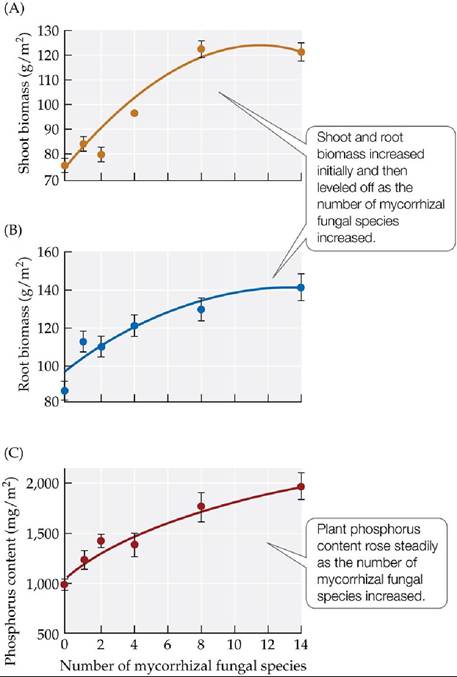
FIGURE 15.20 Mycorrhizal Fungal Species Richness Affects Ecosystem Properties Researchers measured the effects of the number of mycorrhizal fungal species in the soil on (A) mean shoot biomass, (B) mean root biomass, and (C) mean phosphorus content in mixtures of 15 species of plants grown from seed in a field experiment.
Error bars show ± one standard error of the mean. (After M. van der Heijden et al. 1998. Nature 396: 69-72.) View larger imageA Case Study Revisited
The First Farmers
As Figure 15.2A illustrates, the fungal gardens of leaf-cutter ants represent a potentially enormous food resource for any species able to overcome the defenses of the resident ants. In particular, are there any parasites that specialize in attacking fungal gardens? As we saw in Chapter 13, roughly half the world's species are parasites, and many of them have remarkable adaptations for evading host defenses.
Although you might expect that the answer would be yes, for more than 100 years after the fungus-growing role of leaf-cutter ants was discovered (Belt 1874), no such parasites were known. That changed in the early 1990s, when Ignacio Chapela (Seifert et al. 1995) observed that leaf-cutter ant gardens were plagued by a virulent parasitic fungus of the genus Escovopsis (see also Currie et al. 1999a). This parasite can spread from one garden to the next, and it can rapidly destroy the gardens it invades, leading to the death of ant colonies. Leaf-cutter ants respond to Escovopsis by increasing the rate at which they weed their gardens (FIGURE 15.21) and, in some cases, by increasing how often they dose the garden with antimicrobial toxins, which they produce in specialized glands (Fernandez-Marfn et al. 2009).
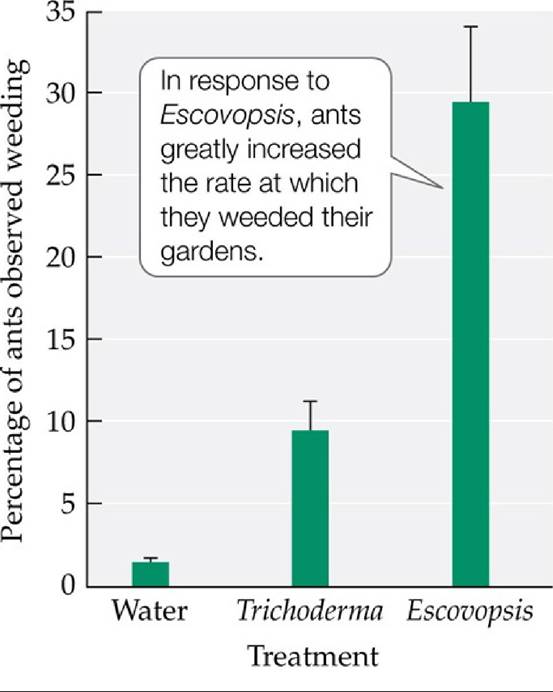
FIGURE 15.21 A Specialized Parasite Stimulates Weeding by Ants Currieand Stuart measured the frequency with which the leaf-cutter ant Atta colombica weeded its fungal gardens after colonies were exposed to water, Trichoderma viride (a generalist fungal parasite), and the specialized fungal parasite Escovopsis. Error bars show one standard error of the mean.
Suppose 2% of ants were observed weeding in colonies exposed to water, 20% in colonies exposed to Trichoderma, and 20% in colonies exposed to Escovopsis.
Propose a hypothesis that might explain these results.(After C. R. Currie and A. E. Stuart. 2001. R Soc 268: 1471.) View larger image
The ants also enlist the help of other species in combatting Escovopsis (Currie et al. 1999b). On the underside of the ant's body lives a bacterium that produces chemicals that inhibit Escovopsis. The queen carries this bacterium on her body when she begins a new colony. While the ants clearly benefit from the use of these fungicides, what of the bacterium? Recent work (Currie et al. 2006) indicates that the bacterium also benefits: the ant provides it with both a place to live (it is housed in specialized structures called crypts that are located on the ant's exoskeleton) and a source of food (glandular secretions). Thus, the bacterium appears to be a third mutualist that benefits from, and contributes to, these unique fungal gardens.
Connections in Nature
From Mandibles to Nutrient Cycling
While you have been reading this chapter, billions of pairs of leafcutter ant mandibles have been removing leaves from the forests of the Americas. The workers of a single colony can harvest as much plant matter each day as it would take to feed a cow. People have long known that leaf-cutter ants are potent herbivores. Weber (1966) describes reports—the earliest from 1559—of leaf-cutter ants destroying the crops of Spanish colonists, and they still plague farmers today. In tropical regions, these ants tend to increase in abundance after a forest is cut down. Anecdotal evidence suggests that the thriving ant colonies found in deforested areas are one of the reasons why farms in some tropical regions are often abandoned just a few years after trees have been removed to make room for them (other reasons relate to a point made in Chapters 3 and 22: some tropical soils are nutrient-poor).
In addition to their effects on human farmers, leaf-cutter ants introduce large amounts of organic matter into tropical forest soils.
As a consequence, they affect the supply and cycling of nutrients in the forest ecosystem (a topic we will discuss in more detail in Concepts 22.2 and 22.3). Normally, nutrients in the leaf litter that falls to the forest floor enter the soil when the leaves decompose. Bruce Haines (1978) compared the amounts (g/m2) of 13 mineral nutrients contained in leaf litter with the amounts of the same nutrients found in aboveground areas where colonies of the leafcutter ant Atta colombica deposit their refuse (other Atta species deposit refuse belowground, as shown in Figure 15.2A). Averaged across the 13 nutrients, the ants' refuse areas contained about 48 times the nutrients found in the leaf litter. Plants respond to this concentration of nutrients by increasing their production of fine roots in the Atta refuse areas. Furthermore, the activities of leaf-cutter ants have the effect of tilling the soil near their nests, making it easier for plant roots to penetrate the soil (Moutinho et al. 2003). Moutinho and colleagues also found that the leaf material ants bring into their colonies fertilizes the soil, causing soils beneath ant colonies to be 3-4 times richer in calcium and 7-14 times richer in potassium than are soils 15 m away from the nest. Finally, recent evidence suggests that the fungal gardens tended by ants may also house nitrogen-fixing bacteria (FIGURE 15.22). These bacteria may be part of yet another mutualism found in the gardens—a mutualism that may prove to be an important source of nitrogen in tropical ecosystems.

FIGURE 15.22 NitrogenFixationinFungalGardens Whenresearchersmeasured nitrogen fixation activity in different parts of the colonies of leaf-cutter ants, they found that most of it was taking place in the fungal gardens. In addition, bacteria from genus Klebsiella were isolated from the fungal gardens and shown to fix nitrogen. Error bars show one standard error of the mean. (After A. A. Pinto-Tomas et al. 2009. Science 326: 1120-1123.) View larger image
The overall effects of leaf-cutter ants on the ecosystems in which they live are complex. In forest ecosystems, net primary production (NPP) is usually measured as new aboveground plant growth (see Concept 20.1); root growth is often ignored, since it is difficult to measure in trees. Although leaf-cutter ants reduce NPP by harvesting leaves, some of the other activities of ants (e.g., tillage, fertilization) may increase NPP. As a result, the net effect of the ants on the NPP of their ecosystem is difficult to estimate. While it may prove possible to disentangle such effects in future studies, there is no doubt that the ants and their partners have considerable effects
on the ecosystems in which they are found.
More on the topic Positive interactions can alter communities and ecosystems:
- CONCEPT 13.5 Parasites can alter the outcomes of species interactions, thereby causing communities to change.
- CONCEPT 15.3 Positive interactions affect the abundances and distributions of populations as well as the structure of ecological communities.
- A general theme that runs through this book is that ecological interactions can affect the distributions and abundances of species, affecting communities and ecosystems.
- Positive interactions may be more common in stressful environments
- Evolution can alter ecological interactions
- Positive interactions can cease to be beneficial under some circumstances
- Carnivores can alter communities in dramatic ways
- Herbivores can alter communities in dramatic ways
- Positive Interactions
- Positive interactions can be obligate or facultative and loosely structured