The equilibrium theory of island biogeography holds true for mainland areas
Do the effects of area and isolation influence differences in species richness in mainland areas as well as on islands? As we saw in Watson's graph of plant species richness in Great Britain (see Figure 18.18), the species-area relationships observed on islands can also hold for mainland areas.
How, then, does the biogeography of mainland areas differ from that of islands and islandlike areas?Let's consider a plot of plant species richness in mainland areas of France and on the Channel Islands in the English Channel (see Ecological Toolkit 18.1). Williams (1964) showed that plant species richness increases with area in both locations but that the slope of the line representing the increase is steeper for the Channel Islands than for the French mainland (i.e., the z value was greater on the islands). How can we interpret this difference? In mainland areas, just as on islands, species richness is theorized to be controlled by rates of immigration and extinction. In mainland areas, however, these rates are likely to be different from those on islands. Immigration rates should be greater in mainland areas because the barriers to dispersal are lower. Species can move from one area to the next, presumably through continuous, non-island habitat. In addition, extinction rates should be much lower in mainland areas because of the continual immigration of new individuals from the larger mainland population. The idea is that species will always have a good chance of being “rescued” from local extinction by other population members. The end result of these higher immigration and lower extinction rates in mainland areas is a lower rate of increase in species richness with increasing area, and thus a gentler slope, than in island areas.
We have seen over and over again in this chapter that geographic area has a large influence on species diversity at global and regional spatial scales.
This effect takes on heightened significance as more habitats become “island-like” because of human influences. As we will see in the Case Study Revisited, the theory and practice of island biogeography is timely and relevant to the issues of conservation that we deal with today.A Case Study Revisited
The Largest Ecological Experiment on Earth
One goal of ecologists is to understand the science behind the conservation of species threatened by habitat destruction and fragmentation. As we set aside more and more reserves to protect species diversity, the areas around those reserves continue to be changed by human activities, leaving many of them islands in a matrix of degraded habitat that is unsuitable for the species they contain. Thus, it is critical that we understand reserve design if we are to meet our conservation goals. When Lovejoy and his colleagues embarked, more than 40 years ago, on the Biological Dynamics of Forest Fragments Project (BDFFP) in the Amazon, one of their goals was to study the effects of reserve design on the maintenance of species diversity (Bierregaard et al. 2001). As it turned out, they learned that habitat fragmentation had even more negative and complicated effects than they had anticipated.
One of the first things they learned was that forest fragments needed to be large and close together to effectively maintain their original species diversity. For example, in a study of forest understory birds, Ferraz et al. (2003) found that even the largest fragments they surveyed (100 ha) lost 50% of their species within 12 years. Given that the regeneration time for these tropical rainforests ranges from several decades to a century, they projected that even fragments of 100 ha would be ineffective at maintaining bird species richness until forest regeneration could “rescue” species surviving within the fragments. The ecologists calculated that over 1,000 ha would be needed to maintain bird species richness until the forests could be regenerated, an area far greater than the average Amazon rainforest fragment in existence today (Gascon et al.
2000). If forest regeneration did not occur—as is likely when the land around a forest fragment is developed or used for agriculture—the fragment would have to contain 10,000 ha or more to maintain most of its bird species over more than 100 years of isolation (although even a fragment of that size could not sustain them all).The researchers of the BDFFP were also surprised at how even minimal distances between fragments resulted in almost complete isolation of species. Clearings even 80 m (265 feet) wide hindered the recolonization of fragments by birds, insects, and arboreal (treedwelling) mammals (Laurance et al. 2002). It seemed that animals avoided entering the clearings for a number of interrelated reasons, the most obvious of which is that they have no innate reason to do so, having evolved within large, continuous, and climatically stable habitats that lacked the fragmentation imposed on them by deforestation. Moreover, even if some animals were inclined to venture into the clearings, specific requirements for their movement, such as trees for arboreal mammals, would not be present to facilitate their travel to other forest patches.
A second major finding of the BDFFP was that habitat fragmentation exposes the species within a fragment to a wide variety of potential hazards, including harsh environmental conditions, fires, hunting, predators, diseases, and invasive species. These edge effects, which occur at the transition between forest and nonforested matrix habitat, can act together to increase local species extinctions. Trees, for example, can be killed or damaged by their sudden exposure to brighter light, higher temperatures, wind, fire, and diseases (FIGURE 18.24). Over time, depending on the surrounding matrix habitat, the ultimate influences of edge effects are revealed. If the matrix habitat is left undisturbed, secondary succession occurs, as described in Chapter 17, reducing edge effects. If the matrix habitat continues to be disturbed, however, then the area subjected to edge effects may increase in size.
For example, Gascon et al. (2000) describe forest fragments in the southern Amazon that are embedded in huge non-native sugarcane and Eucalyptus plantations where burning is used regularly for crop rotation. The burning keeps the forest edges in a constant state of disturbance. Fire-tolerant plant species, many of them non-native, become more common at the edges and act as conduits for more fires. This positive feedback loop ends up decreasing the effective size of the forest fragments and continually increasing the area subjected to edge effects. Some edge effects can extend a kilometer or more into a fragment, essentially influencing the entire area of a 1,000-ha fragment.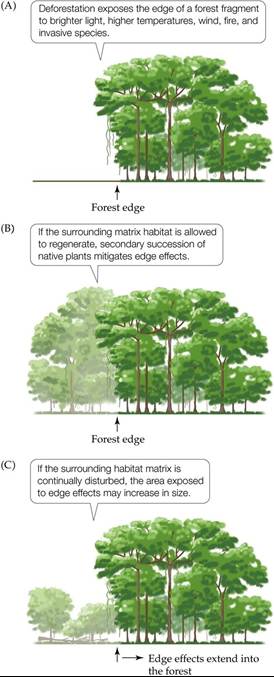
FIGURE 18.24 TropicalRainforestsontheEdge The BDFFP's research showed that deforestation subjects the forest fragments that remain to negative edge effects. (After C. Gascon et al. 2000. Science 288: 1356-1358.) View larger image
The results of the BDFFP have made an immense and sobering contribution to our understanding of forest fragmentation. As Laurance et al. (2002) point out, the BDFFP is a controlled experiment that probably provides a conservative estimate of species losses. The BDFFP has shown us that most of the forest fragments human activities are creating are too small to maintain all their original species; thus, habitat fragmentation is likely to result in the loss of many species. We'll see how the BDFFP's findings are being applied to reserve design and other conservation efforts when we discuss habitat fragmentation and edge effects in more detail in Concepts 24.2 and 24.3.
Connections in Nature
Tropical Rainforest Diversity Benefits Humans
Why do we care when species go extinct in a rainforest far away? As we will see in Concept 23.1, such extinctions raise ethical and aesthetic concerns similar to those that arise when great works of art or antiquities are lost to society.
In addition, there are economic concerns about the loss of important ecosystem services produced by natural systems, which help sustain human health and well-being. For example, tropical deforestation raises concerns about losses of important foods and medicines that have their origins in rainforests. At least 80% of the developed world's diet originated in tropical rainforests, including corn, rice, squash, yams, oranges, coconuts, lemons, tomatoes, and nuts and spices of many kinds. Twenty-five percent of all commercial pharmaceuticals are derived from tropical rainforest plants, but less than 1% of tropical rainforest plants have been tested for their potential medical uses.These statistics raise the question, How does the economic value of tropical rainforest plants used for nontimber purposes compare with the value of deforestation? It turns out that there have been very few economic analyses of this type. A few studies come from the Millennium Ecosystem Assessment (2005), a synthesis of studies on the use of the environment and its relationship to human needs, created by leading scientists from around the world. An example comes from Cambodia, where the total economic value of traditional forest products (e.g., fuelwood, rattan and bamboo, malva nuts, and medicines) was compared with that of unsustainable forest harvesting. The value of traditional forest products is four to five times greater ($700-$3,900 per hectare) than that of unsustainable forest harvesting ($150-$1,100 per hectare).
Recognition of the economic benefits of changing our resource management practices has only just begun. Why is this? Part of the answer lies in our not formally recognizing the economic value of the services provided to humans by species or whole communities. Tropical rainforests provide food, medicine, fuel, and a destination for tourists, all of which can be obtained without complete deforestation. Rainforests also regulate water flow, climate, and atmospheric CO2 concentrations. Assigning a value to any of these important services is difficult compared with setting the market price of timber or agricultural products. For that reason, it is easier to justify the use of rainforest timber and land (and even some sustainable forest products) for private profit than to press for the conservation of rainforests based on the ecological services they provide to society in general. If private landowners are not given incentives to value the larger social benefits of ecological services, maximization of personal gain often drives their decisions. Given the importance of ecological services to our planet, we can no longer afford to ignore these economic trade-offs.