Species richness is a balance between immigration and extinction
The Theory of Island Biogeography (1967) represented one of the most important breakthroughs in the science of biogeography since Wallace's time. The book was born out of the common interests of two scientists: an ecologist, Robert MacArthur, and a taxonomist and biogeographer, Edward O.
Wilson. Wilson, who had studied the biogeography of ants for his PhD thesis work, had made a few key observations about islands in the South Pacific, which he found himself discussing with MacArthur when they met at a scientific meeting (Wilson 1994). The first observation was that for every tenfold increase in island area, there was a rough doubling of ant species richness. The second was that as ant species spread from mainland areas to islands, the new species tended to replace the existing species, but there was no net gain in species richness. There appeared to be an equilibrium number of species on the islands, which was dependent on their size and distance from the mainland, but species composition on the islands could, and did, change over time.MacArthur, a gifted mathematical ecologist, was just 31 years old when he and Wilson developed these observations into the beginnings of a simple but elegant theoretical regional biogeographic model. The model, published in their book 5 years later, became more commonly known as the equilibrium theory of island biogeography. The theory is based on the idea that the number of species on an island, or in an island-like habitat, depends on a balance between immigration or dispersal rates and extinction rates. The theory works something like this: Imagine an empty island open for colonization by species from mainland, or source, populations. As new species arrive on the island, by whatever means necessary, the island starts to fill up. The rate of immigration (the number of new species arriving) decreases over time as more and more species are added, eventually reaching zero when the entire pool of new species that could reach the island and be supported there is exhausted.
But as the number of species on the island increases, there should also be an increase in the rate of extinction. This assumption makes sense according to the simple principle of balance mentioned above: with more species, there are more species extinctions.Additionally, as the number of species increases, the population size of each species should get smaller. Conceivably, this could occur for two reasons. First, competition may increase, thus decreasing the population sizes of species as they vie for the same space and resources. Second, predation may increase as more consumer species are added to the island. The result of either interaction is smaller population sizes and thus a greater risk of species extinction. If we plot the immigration rate against the extinction rate, the actual number of species on the island should fall where the two curves intersect, or where species immigration and extinction are in balance (FIGURE 18.21). This equilibrium number is the number of species that should theoretically “fit” on the island, irrespective of the turnover, or replacement of one species with another, that occurs on the island over time.
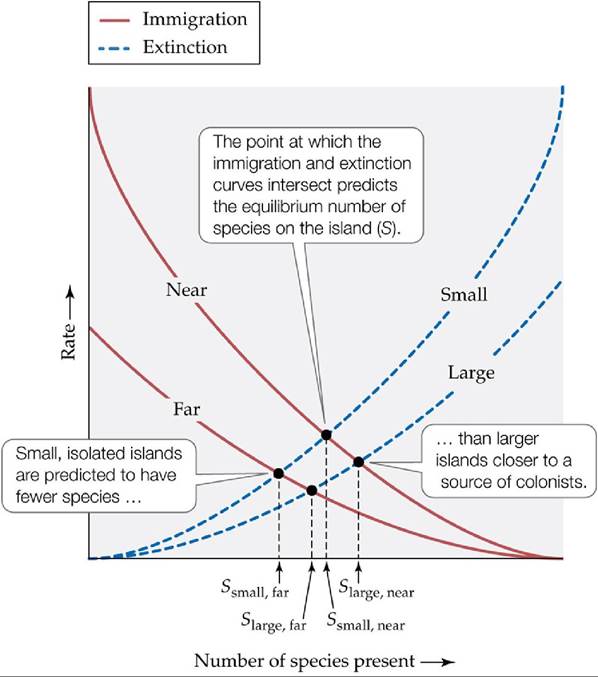
FIGURE 18.21 The Equilibrium Theory of Island Biogeography MacArthur and Wilson's theory emphasized the balance between species immigration rates and species extinction rates for islands of different sizes and at different distances from a source of colonizing species. (After R. H. MacArthur and E. O. Wilson. 1963. Evolution 17: 373-387.) View larger image
To understand the influence of island size and isolation on island species richness, MacArthur and Wilson simply adjusted their curves up or down to reflect their effects (see Figure 18.21). They assumed that island size mainly controls the extinction rate. They reasoned that small islands should have higher extinction rates than large islands, for the same two reasons described above, resulting in an extinction curve for small islands that is higher than that for large islands. Likewise, they reasoned that the distance of an island from the mainland mainly controls the immigration rate.
Distant islands should have lower rates of immigration than islands near the mainland, resulting in an immigration curve for distant islands that is lower than that for islands near the mainland.To test their theory, MacArthur and Wilson (1967) applied it to observations from the small volcanic island of Krakatau, between Sumatra and Java, which erupted violently in 1883, wiping out all life on the island (FIGURE 18.22). Surprisingly, animal and plant species began returning to what little remained of the island within a year of the explosion. MacArthur and Wilson used data from three surveys at various times since the eruption to calculate the immigration and extinction rates of birds on the island. Based on these rates, they predicted that the island should sustain roughly 30 bird species at equilibrium, with a turnover of 1 species. The data showed that bird species richness on the island had indeed reached 30 species within 40 years after the eruption and had remained close to that number thereafter. However, they also found that turnover was much higher, at 5 species. Whether this difference was due to a sampling error or a problem with the model is unknown, but this example motivated Wilson and others (e.g., the BDFFP researchers whose work is described in this chapter's Case Study) to start testing the model using manipulative experiments.

FIGURE 18.22 TheKrakatauTest (A) The eruption of the small volcanic island of Krakatau, near Sumatra and Java, in 1883 provided a natural test of the equilibrium theory of island biogeography. (B) A drawing based on a photo taken from a ship a few hours before the major eruption. (C) By 1921, the number of bird species had reached 31, and in 1934, it was at 30 —the equilibrium number predicted by MacArthur and Wilson's theory. Turnover, however, was five times higher than the theory had predicted. (C after R. H. MacArthur and E.
O. Wilson. 1963.Evolution 17: 373-387; based on data from K. W. Dammerman. 1948. The Fauna of Krakatau: 1883-1933. Noord-Hoiiandsche Uitg.-Mig.) View larger image
One of the best-known experiments to test the equilibrium theory of island biogeography was conducted by Daniel Simberloff and his advisor, Edward O. Wilson, on small mangrove islands and their arthropod inhabitants in the Florida Keys (Simberloff and Wilson 1969; Wilson and Simberloff 1969). These islands were scattered at various distances from large “mainland” mangrove stands (FIGURE 18.23A). After surveying species richness on the islands, Simberloff and Wilson manipulated a handful of them by fumigating them with an insecticide to remove all of their insects and spiders (FIGURE 18.23B). They then surveyed the defaunated islands over a year-long period (FIGURE 18.23C). By the end of the year, species numbers on the islands were similar to those before the defaunation; furthermore, the island closest to a source of colonists had the most species, and the farthest island had the least (FIGURE 18.23D). Interestingly, the farthest island had not quite regained its original species richness even after 2 years. All the islands showed considerable turnover of species, as might be expected for small islands where extinction rates are predicted to be high (see Figure 18.21).
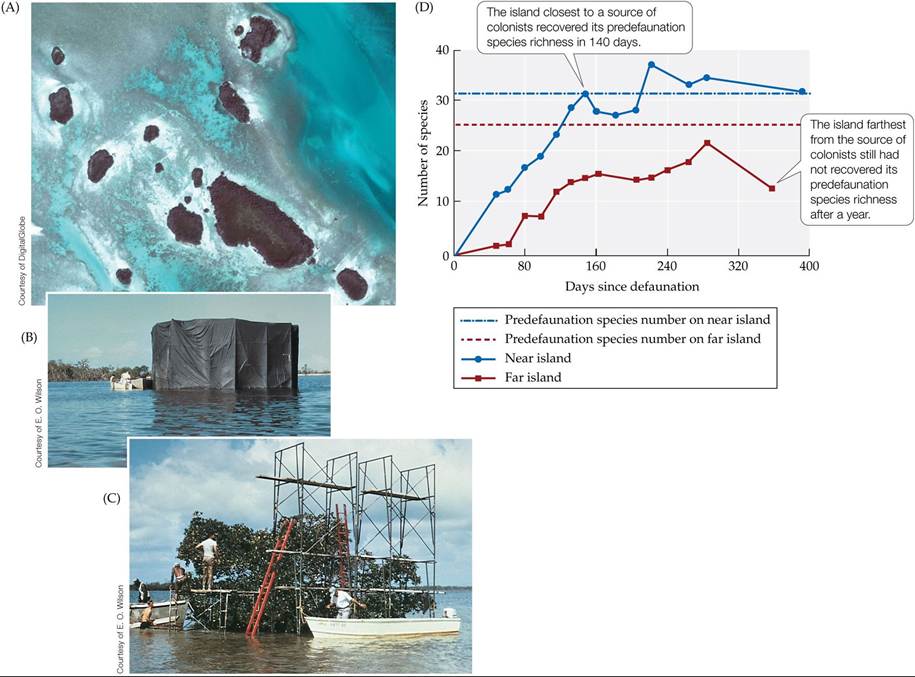
FIGURE 18.23 TheMangroveExperiment (A) To test the equilibrium theory of island biogeography, Simberloff and Wilson surveyed small mangrove islands located at different distances from larger mangrove stands. (B) They then defaunated some of the islands using fumigation tents. (C) They sampled and recorded the number of insect species that recolonized the islands, using scaffolding to reach all parts of the canopy. (D) Results for two islands, one near
and one far from a source of colonists. (D after D. S. Simberloff and E. O. Wilson. 1969. Ecology 50: 278-296.) View larger image