ALTERNATIVE HIV-1 THERAPEUTIC STRATEGIES
Cytokines
Cytokines are soluble, low-molecular-weight, secreted proteins that regulate the amplitude and duration of the inflammatory immune response. They are produced in a transient manner, tightly regulated by the presence of foreign material, and most act locally.
In general, they are pleiotropic, with multiple effects on growth and differentiation of a variety of cell types. There also is considerable redundancy among different types.52 Mossman and Coffman first reported that cloned murine T lymphocytes (CD4+) could be divided into functional subsets on the basis of the immune- regulatory cytokines they produced: Th-1 producing interferon (IFN)-γ,53 Th-2 producing IL-4, and Th-0 producing both.53,54 An inverse relationship was found between cell-mediated immunity (CMI) (mainly sustained by Th-1 cytokines) and humoral antibody (Ab) production (mainly sustained by Th-2 cytokines), with the exception of IgG2a, which is enhanced by Th-1 cytokines.55 This antagonistic relationship between CMI and Ab production may be attributed to the production and crossregulation of Th-1 and Th-2 cytokines. The nomenclature has since evolved to Th-1-like and Th-2- like to reflect the functional plasticity of the two groups and indicate that other cytokines contribute to the regulation of the immune system.56,57 Subsequent studies demonstrated that, in addition to CD4+ T cells, many different cell types produce cytokines, namely, CD8+ T cells, monocytes/macrophages, natural killer (NK) cells, B cells, mast cells, eosinophils, and antigen-presenting cells (APCs). Some researchers also believe that a switch from a Th-1- to Th-2-like immune response may contribute to clinical progression to AIDS.58,59 It was also proposed that Th-1-like cytokines offer some protection from apoptotic cell death, whereas Th-2-like cytokines promote apoptosis of CD4+ cells.60IL-2 and IL-15
Although the mechanism by which IL-2 treatment increases CD4 T cells is not fully understood, it was shown that this increase occurs by the peripheral expansion of existing naive CD4 T cell populations rather than by increased thymic output.61 The intermittent administration of IL-2 to patients receiving continuous HAART was shown to substantially decrease replication-competent HIV in resting CD4 T cells.61 Based on the observation that lymphocytes from HIV-infected individuals express proteins characteristic of G1ZS to G2/M, despite a DNA content and metabolic profile characteristic of the G0 cell-cycle phase, Paiardini and Silvestri et al.
proposed that this cellcycle perturbation may lower the threshold for activation-induced apoptosis.62 They demonstrated that the addition of exogenous IL-2 to HIV-infected lymphocytes induces a profound normalizing effect on the expression of cell-cycle-dependent proteins, thus reducing progression to apoptosis. Data from the same study also indicate that although normal quantities of IL-2 production are observed during HIV infection, its functionality is impaired two- to threefold, and the addition of exogenous IL-2 makes this defect normal.62 This drug, however, should be used with caution in patients with advanced disease, because National Institute of Allergy and Infectious Diseases (NIAID) studies indicate that IL-2 can accelerate disease progression in patients with CD4 counts below 200 cellsZμμ3 by substantially increasing HIV levels.63Both IL-15 and -2 drive T cell proliferation in vitro but are produced by different cell types (IL-2 is produced by activated T cells, and IL-15 is produced by many cell types, including antigenpresenting cells, keratinocytes, and epithelial cells). Proliferation experiments for CD4 and CD8 showed that these subsets exhibit specific sensitivities at different stages of differentiation to IL-2 and IL-15, respectively.64 IL-15 promotes the proliferation and long-term survival of CD8 memory but not CD4 memory T cells in an antigen-independent fashion.
Both IL-2 and -15 upregulate intracellular levels of Bcl-2 and induce the expression of the HIV co-receptor CCR5 (the latter is an important determinant of the level of HIV infection).65 Thus, it is likely that both have discordant (the upregulation of Bcl-2 is a positive feature whereas an increase in the expression of CCR5 may benefit the virus) effects on HIV-infected individuals.66,67 In one study by Zaunders et al.,64 the addition of IL-2 or IL-15 significantly inhibited caspase activation, increased Bcl-2 expression, and rapidly initiated proliferation of CD8 T cells expressing CCR5 in vitro.
Although the authors indicate that inhibition of apoptosis proceeded via the Bcl-2/ cytokine- regulated mitochondrial pathway, they also noted a substantial unexplained upregulation of CD95 expression.64 Another serious consideration with IL-15 administration was raised after one study demonstrated that the expansion of memory cells to concurrent infection in Mycobacterium bovis- IL-15-infected mice coincided with the attrition of existing memory.68 This could obviously have deleterious consequences for the already immune-compromised HIV-1 patient.The production of different cytokines is crucial from a functional point of view and can represent a useful marker to analyze the composition of the pool of virus-specific CD4 T cells. On the basis of the ability to secrete IL-2 and IFN-γ, three functionally distinct populations of CD4 T cells can be identified as follows: IL-2-secreting cells, IL-2∕IFN-γ-secreting cells, and IFN-γ-secreting cells.69 This pivotal study showed that cytomegalovirus (CMV)-specific CD4 T cells were almost equally distributed within the three functionally distinct cell populations. However, a skewing toward IIN- γ-secreting cells (70% of HIV-1-specific CD4 T cells) was observed in subjects with progressive HIV infection, and IL-2-secreting and IL-2∕IFN-γ-secreting cells were almost absent. The frequencies of IL-2-secreting and IL-2∕IFN-γ-secreting HIV-1-specific CD4 T cells were negatively correlated with the levels of viremia. Prolonged antiretroviral therapy was able to correct the skewed representation of different populations of HIV-1-specific CD4 T cells but was associated with only a partial recovery of IL-2-secreting cells. These results underline the importance and role of a decreased capacity to produce IL-2.
IL-12
IL-12 links the innate immune response and adaptive immunity by inducing the early production of IFN-γ by NK cells and later by T cells and is also required for the optimal differentiation of cytotoxic T lymphocytes.70 For these reasons, this cytokine is being used as an adjuvant in several preclinical vaccine trials (mostly cancer studies).71,72 Because vaccine-elicited responses should include long-lived memory cells that can rapidly expand in number following reexposure to antigen, one study was designed to analyze the kinetics, magnitude, and durability of plasmid IL-12- augmented DNA vaccine-elicited responses in mice.73 Mice were immunized with a gp120 vaccine with or without plasmid IL-12 on days 4, 7, 10, and 14.
The authors reported a notable increase in CTL responses in the IL-12 group only at day 10. However, there was no expansion in CTL when boosted with gp120 alone at day 130. Therefore, it is likely that the strong response at day 10 resulted from the expansion of effector memory (instead of central memory) and highlights the importance of timing with cytokine adjuvants.IL-7
IL-7 is another immune-modulating HIV therapeutic candidate. This interleukin provides survival signals to early thymocyte progenitors and protects mature thymocytes from dexamethasone- induced apoptosis,74 which was linked to the upregulation of Bcl-2 in peripheral T cells.75 Unlike IL-2 or phytohaemagglutinin (PHA) treatment, IL-7 can induce the in vitro expansion of naive T cells and maintains their naive phenotype.76 This mechanism was confirmed in the analysis of a small number of chronically infected severely immune-impaired patients who initiated HAART and were capable of reconstituting their pool of CD4 T cells, even in the apparent absence of thymic regeneration.77 Expanded CD4 populations displayed a high percentage of phenotypically naive cells with a very low expression of CD95+, CD45R0+, and CD38+. Plasma levels of IL-7 were significantly high, and most CD4/CD8 cells expressed significant levels of CD-127 (IL-7 receptor) characteristic of healthy donors and had low CD95 levels. There are, however, conflicting reports regarding susceptibility to HIV after IL-7 treatment. The results of one in vitro study indicated that IL-7 promotes both T-tropic and M-tropic HIV replication in naive T cells.76 On the contrary, Napolitano et al. demonstrated in the severe combined immunodeficient with co-engrafted human fetal liver and thymus (SCID-hu Thy/ Liv) mouse model that IL-7 does not increase HIV replication.
Virostatics
A new concept of antiviral/cytostatic (“virostatics”) drugs was proposed within the context of HAART to restrict virus target populations (CD4+ T lymphocytes), target viral reservoirs, and possibly restore immune functions, by reducing excess immune activation.
These virostatics include drugs such as hydroxyurea, mycophenolic acid, and cyclosporin. They use multiple novel mechanisms of action to impede HIV by targeting host cellular proteins that are not susceptible to mutation. Therefore, their resistance profile seems to be favorable.Reducing the hyperactivation of the immune system that accompanies chronic HIV infection might be advantageous. Giorgi et al.10 convincingly showed that immunologic (T cell activation markers) and not virological factors may be more accurate correlates of disease progression. More recently, Silvestri et al. observed minimal immune activation (CD8) and no CD4 T cell depletion in the state of chronically elevated viremia in clinically healthy SIV-infected sooty mangabey monkeys.78 Based on these observations, the authors propose that AIDS progression may involve more than direct virus killing of CD4 T cells. They maintain that excessive turnover rates for both pools of CD4 and CD8 effector cells is most likely the result of AICD6,7,79-83 and constitutes a drain on the naive T cell pool; a reduction in the establishment of long-lived memory cells, which protect the host from secondary infection; and the exaggerated production of effector-generated inflammatory cytokines, which exacerbate immunopathology of HIV and increase viral target populations (CD4 T cells).78
The absence of CD4 T cell depletion despite chronically high levels of virus replication observed in the sooty mangabey model supports the concept that alternative indicators of disease progression may exist. There is substantial evidence that HIV-1 replication requires factors that are made available when host CD4 T cells are activated.84,85 Consistent with this data, it was noted that HIV-1 patients who receive vaccinations against influenza, hepatitis B, or tetanus toxoid (thus, in a state of immune activation) generally experience increases in viral load.84,85 The predator-prey model also substantiates this correlation between the number of available CD4 target cells and virus expansion.
Mathematical models based on the predator-prey equilibrium predict that a sustained expansion of the CD4 T lymphocytes subset—“the prey”—increases the probability that HIV—“the predator”—will spread.86 It was proposed that the cytostatic properties of virostatics may limit the resources made available to the virus, thus keeping their numbers in check.87 Therefore, the quantification and qualitative analysis of CD4 and CD8 T cell subsets and their degree of activation or stimulation may be of clinical interest in predicting progression to AIDS.Although virostatics should function through a common cytostatic mechanism, each needs to be closely evaluated for ulterior immune-modulating properties. For example, both hydroxyurea and mycophenolate acid reduce the nucleotide pool, but each drug acts at a different point in purine synthesis sequence and, thus, there are substantial differences between the two drug mechanisms. Mycophenolic acid (MPA) acts early in the synthesis of purines and has an effect on both transcription and DNA synthesis, whereas hydroxyurea, which interferes with ribonucleotide reductase, primarily effects DNA synthesis (Figure 28.2). Data from recent in vitro and in vivo studies
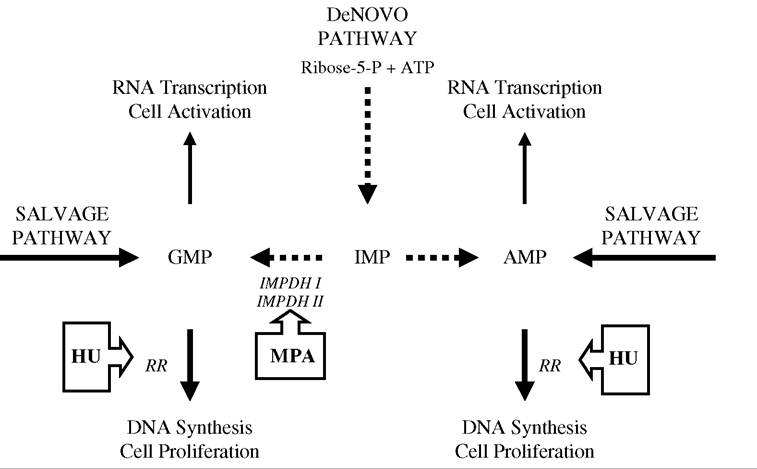
FIGURE 28.2 General synthesis scheme for purines. Two pathways exist for purine synthesis: de novo (dashed lines), mainly used by activated lymphocytes, and salvage (continuous lines), mainly used by resting lymphocytes. Hydroxyurea (HU) inhibits ribonucleotide reductase, thus limiting the production of dNTP synthesis and DNA elongation. Mycophenolic acid (MPA) blocks both RNA transcription and DNA synthesis by targeting the different isoforms of IMPDH (type I and type II). Type I is expressed in resting lymphocytes, whereas type II is primarily expressed in activated lymphocytes and is five times more sensitive to MPA. (See also Konno, Y. et al., J. Biol. Chem., 266, 506, 1991; Nagai, M., Natsumeda, Y., and Weber, G., CancerRes., 52, 258, 1992; Allison, A.C. and Eugui, E.M., Immunopharmacology, 47, 85, 2000.) For these reasons, MPA is expected to be a more potent inhibitor of dNTP synthesis than hydroxyurea in activated lymphocytes, whereas hydroxyurea should be more potent than MPA in quiescent lymphocytes. (RR: ribonucleotide reductase; IMPDH: inosine monophosphate dehydrogenase; IMP: inosine monophosphate; and GMP and AMP are nucleoside precursors.)
clearly indicate that these drugs also effect other changes at the molecular level in various cell types. This is not surprising, because purines not only serve as precursors for DNA/ ribonucleic acid (RNA) synthesis, but they also play an important role in lipid synthesis and protein glycosylation and serve as a source of energy for many other metabolic processes.
Hydroxyurea
Hydroxyurea (hydroxycarbamide) was used in hematology to treat chronic leukemia for more than 40 years and more recently was approved for sickle-cell anemia therapy.88’89 Hydroxyurea uses multiple mechanisms of action to impede the replication of HIV. First, hydroxyurea inhibits the cellular enzyme ribonucleotide reductase, thus blocking the transformation of ribonucleotides into deoxyribonucleotides, depleting the intracellular deoxynucleotide triphosphate (dNTP) pool, and arresting the cell cycle in the G1/S phase,88,89 which directly halts HIV DNA synthesis. Second, synergistic anti-HIV activity was demonstrated when hydroxyurea was combined with NRTIs.90-93 This augmentation in antiretroviral activity may be explained by a favorable change in the proportion of activated NRTI (ddNTP) to dNTP and an enhancement of NRTI phosphorylation.94 Because NRTIs compete with cellular dNTPs for incorporation into the growing HIV DNA, using hydroxyurea to decrease the concentration of cellular dNTP creates a competitive advantage for the NRTIs.95 Consequently, a higher proportion of the NRTI eventually becomes incorporated by reverse transcriptase into viral DNA and thus blocks DNA synthesis. Third, because cell division must progress to the G1b phase of the cell cycle for completion of HIV reverse transcription,96 the virus will only replicate in activated T lymphocytes.97 Thus, when T lymphocytes are treated with hydroxyurea, they remain quiescent and become refractory to productive HIV infection.
Finally, two other interesting, indirect, immune-mediated antiviral characteristics of hydroxyurea were discovered. Heredia et al. found that the arrest of T lymphocytes in the G1 phase of the cell cycle, in response to hydroxyurea, results in the accumulation of -chemokines exerting antiviral activity. These -chemokines, RANTES, MIP-1 and MIP-1 are the natural ligands of the HIV-1 coreceptor CCR5 and compete with R5 HIV-1 virus (CCR5 using strains) for receptor binding.98 In addition, hydroxyurea-containing antiretroviral regimens were shown to restore functional aspects of the immune response impaired by HIV infection, such as reverting the defect of CD3- (the signaling chain of the T cell receptor complex) expression, restoring responses to allogeneic and influenza virus antigens,99 and inducing vigorous HIV-specific T-helper responses.100,101
This drug was extensively evaluated in all three stages of infection (in more than 500 patients) in conjunction with a wide variety of NRTIs. Hydroxyurea was most often paired with didanosine (ddI), a nucleoside analogue, because it augments the activity of this NRTI. Moreover, even though this combination does not prevent the emergence of mutant viral strains, it renders these strains sensitive to ddI.92,102 However, due to the potential for hydroxyurea to increase the toxicity of certain NRTI analogues, a dosing study of the drug was warranted. RIGHT 702 was a randomized, controlled, hydroxyurea dose-regimen study of 115 HIV-infected patients, to compare safety and efficacy of hydroxyurea administered at three daily doses (600, 800 to 900, or 1200 mg/day) and at three dosing intervals—qd, bid, or three times daily (tid)—in combination with ddI and stavudine (d4T). The primary objectives of this study were to determine the safety and tolerability of hydroxyurea administered at different daily doses as well as compare antiretroviral activity at these same doses in combination with ddI and d4T. This study provided the most convincing data supporting the clinical value of an appropriately dosed hydroxyurea/ddI regimen. Surprisingly, the lowest 600-mg dose in the RIGHT 702 study proved the most efficacious in suppressing viremia and was associated with fewer adverse events and the highest CD4 count increase.
Our laboratory recently showed in an in vitro analysis of T lymphocyte proliferation that hydroxyurea has no relevant effect on the expression of activation markers. Moreover, this drug was found to increase the number of apoptotic cells as visualized with Annexin V staining (see MPA section) only at concentrations above 10 μL.103 Considering that in vivo pharmacokinetic analyses indicate that a 1000-mg total daily dose corresponds with an in vitro dosage of 135 μL, and the drug demonstrates linear pharmacokinetics (absorption and elimination),104 a lower 600-mg total daily dose should only minimally induce apoptosis. This, of course, needs to be confirmed with new in vivo pharmacokinetic studies at this concentration.
Mycophenolate Acid
MPA is another interesting candidate for the treatment of HIV/AIDS. Mycophenolate mofetil (CellCeptTM), the prodrug of MPA, is licensed for the prevention of renal transplant rejection and was extensively investigated in different models of allogeneic transplantation, autoimmune skin disorders, and rheumatoid arthritis.105-107 Its principal mechanism of action is the inhibition of inosine monophosphate dehydrogenase (IMPDH), which is the rate-limiting enzyme in the de novo synthesis of guanosine nucleotides. Because MPA selectively inhibits synthesis of guanosine nucleotides, it potentiates the antiviral activity of guanosine inhibitors such as abacavir. There also exist different isoforms of IMPDH (type I and type II): type I is expressed in resting lymphocytes, whereas type II is primarily expressed in activated lymphocytes and is five times more sensitive to MPA.108-110 Based on this information, MPA should limit cellular proliferation as well as the expansion of viruses in activated T lymphocytes but likely has no effect on latently infected resting cells (Figure 28.1).
One common method for investigating apoptosis is to look at the phospholipid phosphatidylserine, which is translocated to the outer plasma membrane during the early stages of apoptosis. This molecule will bind the marker Annexin V, which may be visualized and quantified by FACS analysis. Chapuis and Pantaleo et al., using this system, found that MPA suppresses proliferation in the presence of IL-2 by inducing apotosis in activated, but not resting, T cells.111
Cyclosporin
Cyclosporin is a known immunosuppressant that is used extensively to prevent organ transplantation rejection and was proposed to have a possible role in HIV treatment. Cyclosporin forms a complex with cyclophilin A, which inhibits the calcium and calmoduline-dependent protein phosphatase calcineurin, thus ultimately blocking the activation of genes for IL-2, -4, and the -2 receptor in T cells.112 Rizzardi et al. maintain “that shutdown of T cell activation (with a drug such as cyclosporin) in the early phases of primary HIV-1 infection can have long-term beneficial effects and establish a more favorable immunologic set-point.”112,113 Regarding apoptosis, one recent study with anti-CD3/anti-CD28-stimulated T lymphocytes from healthy donors demonstrated with Annexin V staining that cyclosporin inhibits AICD.114 This drug also directly acts against HIV by preventing proper HIV virion mutation. Finally, it was postulated that cyclosporin synergizes with protease inhibitors by augmenting their absorption. However, due to this drug’s intrinsic immunosuppressive effects (blocking the activation of genes for IL-2, the -2 receptor, and -4), it should be scrutinized to test whether it could become a candidate for HIV treatment.