GP120 RECEPTOR BINDING
CD4 Binding
The HIV-envelope gp120 must bind two receptors in order to enter and infect a host immune cell. The gp120 binds to the CD4 receptor as well as a chemokine co-receptor, either CCR5 or CXCR4, depending on whether the viral strain is R5 or X4, respectively (Figure 6.1).
Numerous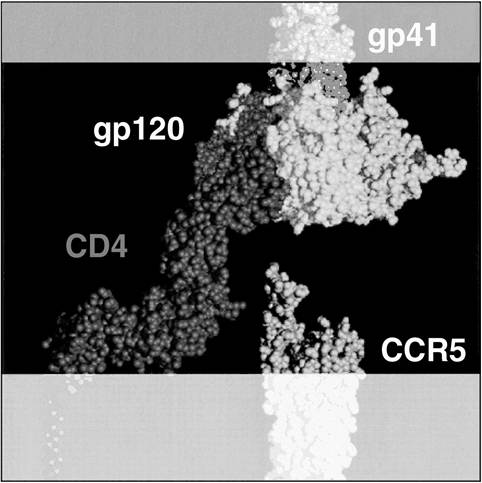
FIGURE 6.1 Structure of HIV envelope and host cell receptors. X-ray crystallographic adaptation showing HIV envelope gp120 and gp41 in proximity with the host cell receptors CD4 and chemokine co-receptor CCR5.
other G-protein-coupled chemokine receptors have been shown to function in vitro as co-receptors for HIV-1, HIV-2, or SIV. However, there is little evidence to suggest that any of these alternative co-receptors have a role for HIV binding or signaling in vivo.21
CD4 is the primary receptor for HIV binding and cell entry.22 CD4 is located on the surface of many helper/inducer T cells, monocytes, and macrophages. The CD4 is a 55 kDa glycoprotein with an N-terminal extracellular domain, one transmembrane segment, and a C-terminal intracellular tail. The extracellular portion includes residues 1 to 37 that form four immunoglobulin-like domains (D1 to D4) that can be recombinantly expressed as soluble CD4 (sCD4).23 X-ray crystallography studies of a CD4/gp120/neutralizing antibody complex have defined the exact location and nature of the HIV gp120/CD4 interaction. The gp120 binds to the D1 and D2 fragments of the CD4 receptor. CD4 is bound to gp120, with the N-terminal domain of CD4 being rigidly held in a deep-binding pocket of gp120.23 Neither the variable loops of gp120 nor any of the carbohydrate moieties near the binding site for CD4 interfere with the orientation of binding of either partner. Direct contact is made between 22 CD4 residues and 26 gp120 residues, comprising, in total, 219 van der Waals contacts and 12 hydrogen bonds.
Chemokine Receptor Binding
CXCR4 and CCR5 are members of the chemokine receptor family. They have seven transmembrane domains and are G-protein coupled receptors (GPCRs). Their functions can vary widely according to their microenvironments, possible receptor dimerization, and posttranslational modifications. The posttranslational modifications of CCR5 and CXCR4 include N-linked and O-linked glycosylation and tyrosine sulfation of the extracellular domains of the receptors, such as N-terminal, extracellular loop I (ECI), ECII, and ECIII. The intracellular loops of the receptors undergo palmtoylation, phosphorylation, and ubiquitination. Each of these modifications can play a role in receptor internalization and receptor turnover.24
After the gp120 binds to the CD4 receptor, it undergoes a conformational change in structure, which better exposes the residues on the envelope glycoprotein and thereby improves binding to the appropriate chemokine co-receptor. A large amount of work has been done to decipher the critical areas of the co-receptors for gp120 binding. This work has demonstrated that the N-terminal and second extracellular domain (ECD) of the co-receptor are crucial for gp120 binding and fusion. Although the interaction of HIV within these regions varies among strains, all the Cys residues on the chemokine receptors seem to be important for co-receptor function, suggesting that these residues are responsible for disulfide bonds. Deglycosylated chemokine receptors are still expressed normally on the cell surface and can still function as co-receptors, suggesting that the sugar moieties are not required for gp120 binding.25 In fact, a deglycosylated CXCR4 has expanded tropism and can bind to dual and some R5 tropic viruses, as well as X4 viruses.26
Several factors determine how likely the host immune cell receptors will bind to gp120 and, therefore, allow entry and infection of the cell. The density of the CD4 receptors on the cell surface as well as CCR5 and CXCR4 conformations influences envelope binding and may explain differences in infectivity between cell types.27 There are a number of posttranslational modifications to both chemokine co-receptors that alter their function by changing their surface density and thereby affecting the affinity of the gp120/co-receptor interactions.
The carbohydrate moieties expressed by the receptors control stability, intracellular trafficking, surface expression, and protein folding. The glycosylation sites on seven-transmembrane domain receptors are generally at the N-terminus sequence. CXCR4 contains two potential sites for N-linked glycosylation: one at the N-terminus and one in the ECII loop.28 Removal of one or both glycosylation sites from CXCR4 results in an expanded tropism of the receptor. With both sites gone, the receptor can recognize and bind several R5 strains of HIV. The CCR5 receptor has only one potential N-linked glycosylation site, but it is not used. In multiple cell lines as well as primary macrophages, the CCR5 receptor has O-linked glycosylation at the N-terminus. Removing these sites does not alter HIV entry but blocks the binding of its natural chemokine ligands MIP-1α and MIP-1β.29
Tyrosine sulfation is another posttranslational modification that is common to many proteins and is often found in tyrosine-rich regions of proteins such as the N-terminal regions of CCR5 and CXCR4. Preventing tyrosine sulfation on the chemokine co-receptors significantly decreases HIV entry but does not alter expression of the receptor on the cell surface.30 However, the studies of tyrosine sulfation were performed in cell lines with transfected co-receptors, making it difficult to determine the physiological role of sulfation modifications and their impact on HIV infectivity in the host immune cell. It is clear that glycosylation and sulfation of the chemokine co-receptors’ extracellular domains alter the overall negative charges of the receptors and thereby their specificity for binding gp120.
CCR5 and CXCR4 are internalized and recycle back to the cell surface. Multiple studies using truncated and mutated forms of the co-receptors demonstrate that receptor internalization is not required for HIV entry into the cell. The receptor turnover rate, however, affects the concentration of receptors on the cell surface that are available for viral binding. G-protein-coupled receptors internalize via agonist-induced endocytosis in clathrin-coated pits. After internalization, GPCRs follow one of two pathways. They either recycle back to the cell surface, or they move to lysosomes for degradation.31 An immediate serine phosphorylation of the cytoplasmic tail of CXCR4 and CCR5 is needed for receptor internalization. This function seems to be modulated by GPCR kinases or protein kinase C.32,33