IMPACT OF HIV INFECTION ON THYMIC FUNCTION
Alteration of Thymic Output after HIV Infection
During HIV infection, de novo T cell production by the thymus is critical for immune reconstitution, as it ensures the replenishment of the T cell pool with a functionally diverse TCR repertoire.
Patients infected by protease inhibitor (PI)-resistant HIV strains with diminished capacity to infect thymocytes maintain normal CD4 T cell counts despite high viremia.32 Many studies have demonstrated the deleterious effect of HIV on thymopoiesis. Thymic specimens from HIV-infected individuals exhibit a profound disorganization of the thymic stroma associated with depletion of thymocytes.33,34 Immunohistochemical studies in these tissues as well as in the SCID-hu Thy/Liv mice (SCID mice with a human thymus grafted on the renal capsule) further demonstrated that HIV-infected thymic tissue contained RNA and viral proteins within the resident thymocytes, confirming their susceptibility to infection.35-37Early reports indirectly suggested a decrease in thymic output, as the frequency of naive T cell (CD45RA+) in the peripheral blood was found to drop with HIV disease progression.38-40 However, this decrease in naive T cell numbers could be the result of increased peripheral T cell activation, a hallmark of HIV infection. HIV-infected individuals consistently show the upregulation of several activation markers, such as CD38, HLA-DR, and Ki67, reflecting their heightened immune activation and the increased recruitment of naive cells to the effector memory compartment.41-44 Recently, quantification of the frequency of T cell receptor excision circles (TRECs), which are markers for intrathymic TCR rearrangements, has led to much insight on thymic function in HIV- infected patients. Many studies demonstrate the reduced frequency of TRECs in peripheral blood mononuclear cells (PBMCs) and purified naive T cells in untreated, HIV-infected individuals.45-48 A large-scale study (297 HIV-infected patients who ranged in age from 20 to 59 years) using sorted CD4 and CD8 T cells demonstrated that TREC frequency increases in CD4 T cells early during HIV infection and slowly declines, whereas CD8 T cells harbor much lower TRECs throughout disease progression.49 Longitudinal analysis of SIV-infected macaques also showed that TREC levels decline in CD4 and CD8 T cells at 16 to 30 weeks after infection, which correlates with an increase in CD8 T cell infiltrates and apoptosis in the thymus, linking affected thymic function and reduced TREC frequencies.50 Finally, in contrast to lower TREC levels in rapid-progressing HIV- infected children, TREC levels in long-term asymptomatic, vertically HIV-infected children are similar to TREC levels in uninfected individuals.51 Again, this stresses the impact of thymic output in maintaining CD4 T cell count after HIV infection.
However, such observations must be interpreted cautiously given the higher proliferation rate of PBMCs within HIV-infected patients, which could also result in the decrease in TREC numbers. The exact contribution of thymic output or peripheral proliferation to the reduced TREC frequency remains to be assessed.52,53 Interestingly, highly active antiretroviral therapy (HAART) was shown to trigger an increase in TREC frequency to levels comparable to age-matched controls in HIV- infected children and adults.54-56 This was, in most cases, accompanied by an increase in thymic size (as determined by computerized tomography [CT] scan),57 suggesting an improved thymic function when HIV viral load was controlled. But again, the increase in TREC levels is also accompanied by a decrease in peripheral T cell activation and, thus, a diminished TREC dilution effect.43,58 Still, the observed increase in TCRβ variable region diversity within CD45RA+ CD8+ T cells in HIV-infected children after HAART suggests a replenishment of the T cell repertoire with new T cells from the thymus and, thus, an increase in thymic function.59 Taken together, these data suggest that thymic function seems to be reestablished following suppression of viral load, which correlates with observations demonstrating the cytopathic effect of thymocyte infection by HIV.
Several Thymocyte Subsets Are Susceptible to HIV Infection
Most thymocytes, including the CD3- CD4+ CD8- cells (immature single-positive [ISP]), CD3+ CD4+ CD8+ (DP cells), and CD3+ CD4+ CD8- single-positive (SP) cells, carry the CD4 molecule and are thus a potential breeding ground for the HIV virus. However, various thymocyte subsets exhibit a different sensitivity to HIV infection, and this mainly depends on their levels of HIV coreceptor expression (Figure 20.3).
Although CXCR4 expression is high in DN, ISP, and DP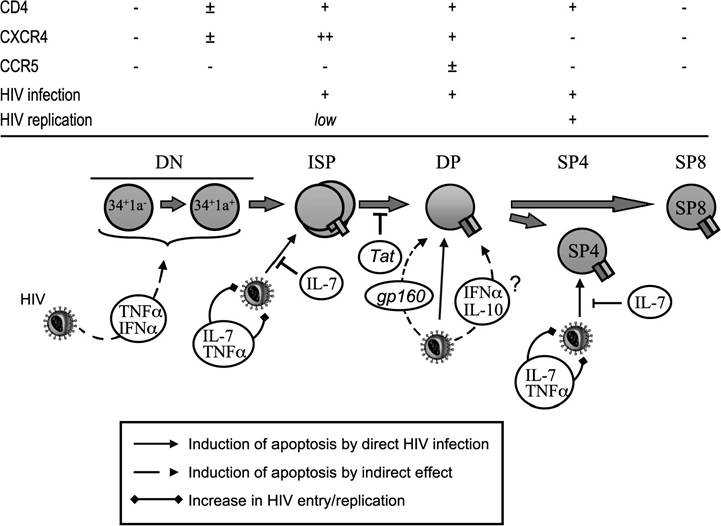
FIGURE 20.3 Influence of HIV on human thymic development. HIV infection directly causes an increased apoptotic rate of immature single-positive (ISP), DP, or CD4+ SP thymocytes (plain arrows). Thymocyte maturation is also indirectly altered by the increase in cytokine levels or the presence of HIV proteins (dotted arrows). An increase in TNF-α or IL-7 levels enhances HIV replication in thymocytes and the pathogenic effect of HIV on thymic function. The levels of CD4 or HIV co-receptor expression are indicated for each subset, as well as the susceptibility to infection and the capacity to sustain HIV replication.
thymocytes, it decreases considerably in mature SP thymocytes.60-62 On the contrary, CCR5 expression is very low throughout the differentiation process and is restricted to 1 to 3% of thymocytes.61-63 Correlating with these observations, CXCR4-using viruses (X4) have a stronger cytopathic effect on thymocytes than the CCR5-using strains (R5) in the SCID-hu mouse model61 or in SIV-infected infant macaques.64 In the SCID-hu model, thymic dysfunction was mainly attributed to a preferential infection of ISP by HIV.65 Furthermore, SIV infection seems to inhibit the developmental potential of ISP; when exposed to SIV, these cells differentiate less in a xenogeneic monkey-mouse fetal thymus organ culture (FTOC) system than ISP from uninfected monkeys.66
Experiments using mixed thymocyte-stromal cells co-cultures have demonstrated that even though HIV can efficiently enter ISP and DP cells, virus production occurs only in SP CD4+ cells.67 Although this subpopulation possesses a low level of CXCR4 expression, exposure to interleukin (IL)-7, a cytokine secreted by thymic epithelial cells (TECs), upregulates the CXCR4 molecule to the cell surface, rendering them susceptible to X4 HIV-1 entry.68 Furthermore, it has been shown that IL-7 regulates TNF-α expression, which, in turn, induces the activation of the NF-κB transcription factor, required for high replication of HIV.67,69 Finally, although CD8 SP thymocytes are not directly susceptible to HIV infection, HIV provirus has been detected in this subset. Infection of DP thymocyte has been shown to produce CD4- CD8+ SP cells that contain HIV provirus, which could contribute to the latent HIV reservoir, because after stimulation in the periphery, these cells can sustain HIV replication.70,71 In summary, several thymocyte subsets are susceptible to HIV infection (ISP and DP) and are able to sustain replication (SP). The evidence that HIV infection results in the apoptosis of some of these subsets will now be described.