ROLE OF APOPTOSIS IN T CELL DEVELOPMENT
In more than 95% of cases, the fate of a developing T cell in the thymus is death; thus, apoptosis is a routine process within the thymus. Two main options can lead a thymocyte to programmed cell death: either the lack of TCR signaling, termed “death by neglect,” or a signal that is too strong, termed “negative selection,” both of which contribute to maintaining a functional yet not autoreactive T cell pool.
The remaining 5% of thymocytes able to engage a major histocompatibility complex (MHC)-peptide complex with relatively weak affinity are positively selected and survive the maturation process to be released in the periphery first as recent thymic emigrants (RTEs), which later become mature T cells (Figure 20.2).16Death by Neglect
During their maturation, thymocytes rearrange the V(D)J segments of the TCR beta (TCRB) locus followed by the TCR alpha (TCRA) locus, leading to the production of T cells with a diverse repertoire of TCRs. Such a random process evidently will yield a large number of “out of frame” rearrangements as well as TCRs that are unable to bind to the MHC-peptide complex or even remain on the surface of the cell, thus rendering many thymocytes unfit to continue differentiation. These thymocytes undergo apoptosis through death by neglect, due to a lack of signal either from the pre-TCR (pTα-β) or from the TCR (α-β). In mice that cannot rearrange the β chain (Rag1-/-, Rag2-/-, and severe combined immunodeficient [SCID] mice) that leads to the absence of pre-TCR signaling, there is a developmental arrest and apoptosis at the CD4- CD8- CD25+ CD44- pro-T3 stage (where the pre-TCR is usually expressed).17,18 DR signaling and the extrinsic pathway may play a role in the depletion of these thymocytes.
Although Fas deficiency (lpr mice) does not increase the survival of thymocytes lacking the pre-TCR to the DP stage, deletion of FADD or expression of a dominant negative mutant of FADD permits their survival and differentiation.19,20 This suggests that a DR other than Fas is involved in death by neglect. Cell death due to the absence of pre-TCR signaling may also involve the tumor suppressor p53 and the intrinsic pathway, because RAG-deficient or SCID mice lacking p53 contain thymocytes past the pro-T3 stage.21-23 Moreover, many reports have underlined the importance of the intrinsic pathway, modulated by the Bcl-2 family proteins, in death by neglect of DP thymocytes. Among them, proapoptotic Bax and Bak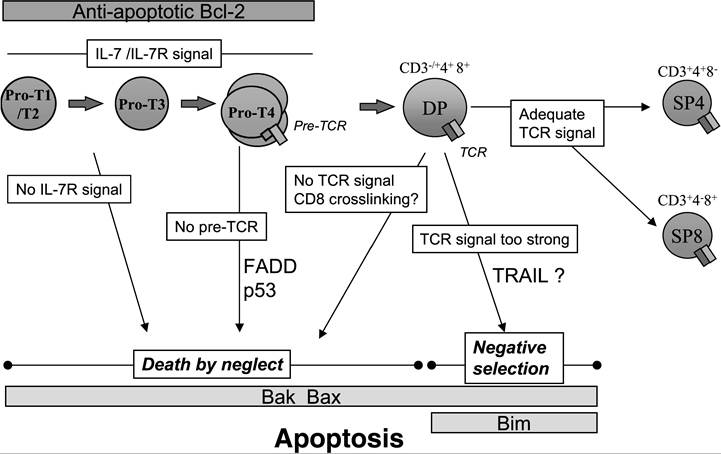
FIGURE 20.2 T cell development and apoptosis in the murine thymus. Double-negative thymocytes (pro-T1 to pro-T4) that do not express a functional TCR undergo apoptosis by a process called “death by neglect.” Once they mature into CD4+ CD8+ double-positive (DP) thymocytes and express the TCR, the absence of TCR signaling also results in death by neglect. Engagement of the TCR with a weak affinity triggers DP cell survival and differentiation into single-positive (SP) mature thymocytes (positive selection), whereas a high-affinity interaction with the MHC-peptide complex results in the deletion of the T cell clone by negative selection.
seem to play a role in the induction of cell death by triggering the disruption of mitochondrial transmembrane potential in “neglected” thymocytes. Thymuses reconstituted with Bak-/- Bax-/- hematopoietic cells display an accumulation of CD4- CD8- thymocytes that do not differentiate into DP cells. These thymocytes do not die by neglect, as they maintain a constant viable cell count for 7 days as in vitro culture in medium alone.24 Therefore, signals leading to the mitochondria are required for the induction of death by neglect.
After TCR rearrangement produces a functional receptor, TCR engagement on DP thymocytes can provide either a rescue signal, which counteracts death by neglect (positive selection), or a death signal (negative selection).Negative Selection
Negative selection is triggered after high-affinity TCR engagement by MHC-self-peptide complexes. The role of DR in this process has been extensively studied, and most studies suggest that DRs are not required for negative selection. Mice expressing a FADD dominant negative form, which inhibits signaling though Fas, TNFR1, DR3, and TRAILR2, exhibit normal or even increased negative selection compared with their wild-type counterparts, again suggesting a possible prosurvival role of FADD.25 However, the importance of TRAIL in negative selection recently became a matter of debate. An impaired deletion of thymocytes by TCR ligation was initially observed in TRAILdeficient mice.26 Contrarily, using various “thymic selection” models such as antibody-mediated TCR cross-linking, stimulation with endogenous superantigens, or exposure to exogenous superantigen in vitro, another group could not identify a defect in negative selection in thymocytes from TRAIL-/- mice.27 Thus, the role of TRAIL in negative selection remains to be confirmed.
In contrast, the intrinsic pathway plays a critical role during negative selection, although the antiapoptotic protein Bcl-2 is not involved. When overexpressed, Bcl-2 protects immature thymocytes from selective death stimuli in vitro, including glucocorticoids, irradiation, and TCR cross-linking. However, in vivo, Bcl-2 overexpression only modestly reduces negative selection mediated by histocompatibility (HY) antigen28 or endogenous superantigens.7 On the other hand, the proapoptotic BH3-only Bcl-2 family member Bim is essential for negative selection, as Bim-deficient transgenic thymocytes expressing a HY-specific TCR are resistant to apoptosis in the presence of the HY negative-selecting peptide.29 Moreover, ligation of the TCR induces the accumulation of Bim and its association with Bcl-xL,29 confirming its involvement during negative selection.
Bax and Bak are also important players in thymocyte deletion, as negative selection is impaired in thymocytes from Bax and Bak double knockout mice.24 These observations support a critical role of mitochondria in the control of apoptosis during negative selection, although Bcl-2 does not seem to be involved. The downstream events leading to cell death are not so clear. Mice deficient in either apoptotic activating factor 1 (Apaf-1) or caspase-9 do not show abnormalities in thymocyte selection,30,31 suggesting that formation of the apoptosome is not essential during negative selection.In summary, a large number of studies have focused on the molecular pathways involved in the deletion of immature thymocytes in vivo by death by neglect or negative selection. Although the survival of T cell progenitors is without doubt controlled by the mitochondria, the exact mechanisms involved in thymocyte deletion in the thymus still need to be clarified.