INTRODUCTION
The Nef protein of the human immunodeficiency virus (HIV) is as important for HIV disease progression as it is perplexing in its number of target molecules and functions. In the early years of acquired immunodeficiency syndrome (AIDS) and Nef research, Nef was thought to be a negative factor (Nef for negative), moderately repressing HIV transcription.1 This view changed completely in 1991, when Kestler and colleagues reported that Nef is essential for the maintenance of highly viral loads and, thus, progression to AIDS in the rhesus macaque animal model.2 Their finding was subsequently confirmed in numerous reports using animal models and was corroborated by follow-up studies of individuals who were infected with Nef-defective viruses but did not progress in the disease.3,4 Today, it is generally accepted that Nef quite drastically increases the viral load, but, surprisingly, after almost two decades of research, the underlying molecular mechanisms that would explain that phenomenon are still not understood.
Nevertheless, there seems to be a consensus that several effects and functions of Nef contribute to the overall increase of viral load in the infected host. From a molecular point of view, Nef has two functions. First, it stimulates T cell activation pathways, and second, it interferes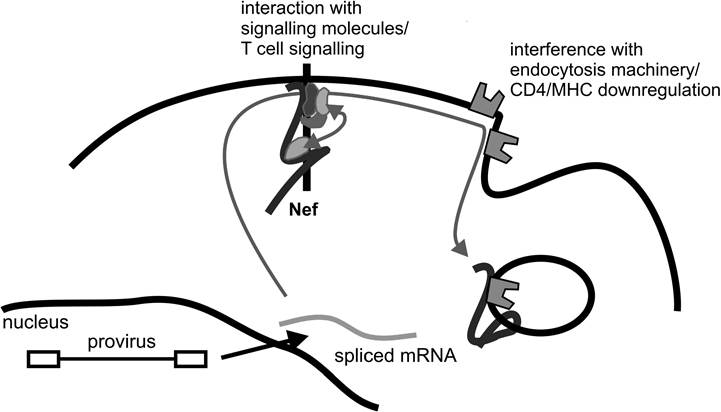
FIGURE 8.1 The two molecular functions of Nef. Nef is expressed early in the viral life cycle from spliced messenger RNA. After protein translation, the protein is translocated to the plasma membrane. There, upon anchoring into the membrane via the myristoylated N-terminus, possibly a conformational change occurs, enabling the protein to associate with signaling molecules from the T cell receptor environment and leading to the assembly of the Nef signaling complex.
Subsequently, Nef interacts or binds cell surface receptors like CD4 and MHC class I, causing their internalization and degradation.with cellular trafficking and the endocytosis machinery (Figure 8.1). Surprisingly, these molecular functions of Nef do not seem to stimulate viral replication directly. Rather, particle production and release from infected cells are increased by indirect means. Despite circumstantial evidence that Nef expression might stimulate replication directly, it has never been formally proven.
Additional indirect effects of Nef are mechanisms that both induce and block cellular death programs. At first glance, it seems to be contradictory that one protein induces opposite effects to support one function, which is the increase of viral particle production. However, as will be discussed below, these effects may not occur at the same time in the viral life cycle and are, therefore, not mutually exclusive. Interestingly, both molecular functions of Nef, T cell signaling and interference with endocytosis, converge to manipulate cellular death programs. The co-evolution of two obviously different molecular mechanisms in one small viral protein is remarkable and again demonstrates the complexity and sophistication of HIV and simian immunodeficiency virus (SIV).
It is generally believed that the increase of viral load is the only contribution of Nef to disease progression. In the mouse model, however, Nef expression alone is sufficient to induce an AIDSlike disease with T cell depletion and immunodeficiency.5 Although there are many differences between transgenic expression of Nef in mice and natural infection of humans and monkeys, it cannot be ruled out that Nef has direct detrimental effects on the immune system in the absence of any viral replication. Such effects, however, have not yet been sufficiently described.