TCR SIGNALS THAT STIMULATE ANTI- AND Proapoptotic pathways
The T cell receptor is one of the best studied of the cellular surface receptors. When pro- and antiapoptotic signaling pathways were detected and described, it was soon learned that this receptor is not only a key regulator of proliferation but also of cellular death programs in T cells.38 This is because both T cell activation/cellular proliferation and T cell death are intimately correlated.
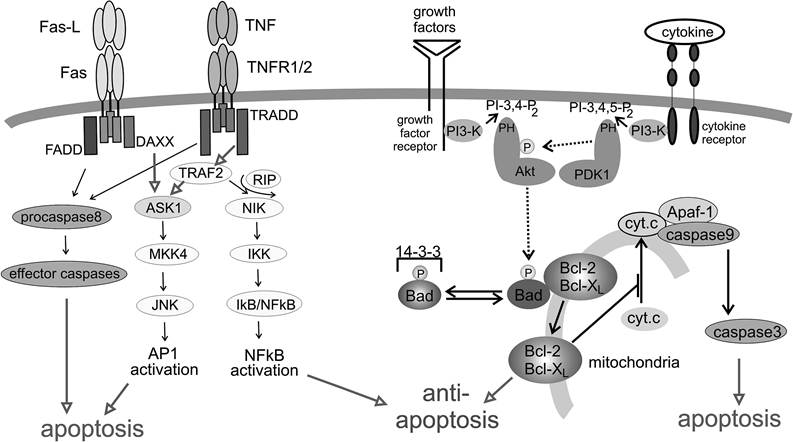
FIGURE 8.2 The “intrinsic” and “extrinsic” apoptotic pathways. Apoptotic cell death is principally induced by two different molecular mechanisms. The extrinsic pathway transmits extracellular signals through death receptors such as Fas and TNFR, as depicted on the left side of the figure. The intrinsic pathway is activated through internal sensors (for example, p53, not shown), leading to the release of cytochrome c from mitochondria and subsequent activation of caspases (right lower part of the figure). The release of cytochrome c is regulated through members of the Bcl-2 family, which again are under the influence of signaling pathways emanating from surface receptor and PI3 kinase (upper right part of the figure).
In recent years, the PI3 kinase-Akt signaling pathways (Figure 8.2) have emerged as an important and general mechanism of antiapoptosis.39 In T cells, this pathway is activated through stimulation, for example, through the T cell receptor or growth factor receptors. Beyond that, the constitutive upregulation of PI3 kinase-Akt survival signaling has also been implicated in oncogenic transformation and tumor development, as it prevents apoptotic cell death during uncontrolled proliferation. The phosphatidylinositol-3 kinases (PI3 kinases) comprise a family of heterodimeric proteins that produce lipid second messengers by phosphorylation of plasma membrane phosphoinositides at the 3 OH of the inositol ring.
The products of this enzymatic reaction (3,4-biphosphate and 3,4,5-triphosphate) are anchored at the plasma membrane and subsequently recruit cellular signaling molecules that contain a lipid binding — a so-called pleckstrin homology (PH) domain.40 PI3 kinase is activated through the T cell receptor or CD28. A precise mechanism is not clear but likely involves other proteins, such as the tyrosine kinase Lck and the adapter proteins Gab 1 and 2.38 A wide variety of signal transduction proteins interacts with PI3 kinase products, one of which is the Akt kinase. Akt was identified as an enzyme similar in many respects to protein kinase A (PKA) and protein kinase C (PKC), and it is, therefore, also referred to as protein kinase B (PKB). After it is recruited to the plasma membrane, Akt is activated by phosphorylation at threonine 308 by another PH-domain-encoding kinase, PDK-1. After its activation, Akt phosphorylates a number of cellular signaling proteins, including the proapoptotic Bad protein (Figure 8.2), a member of the Bcl-2 family that regulates cell death through mitochondria-associated proteins.41 Akt-phosphorylated Bad is inactivated through association with 14-3-3 proteins, which prevents Bad from dimerizing with and inactivating pro-survival members of the same family, for example, Bcl-2. However, in T cells, this mechanism or pathway may not play a major role in survival, as Akt transgenic T cells do not display increased phosphorylation of Bad.42 Rather, it has been suggested that Akt activation leads to the activation of NF-kB, which, in turn, would lead to transcriptional upregulation of Bcl-XL that protects the cell from proapoptotic effects of Bad (also see discussion below). Potential mechanisms of Akt-mediated NFkB activation are controversial and are not understood at this time.In addition to antiapoptotic signals, the T cell receptor regulates proapoptotic signals through activation-induced cell death (AICD), a mechanism that is thought to limit clonal expansion of T cells and cause the deletion of T cell clones.43 The latter mechanism is particularly important in protecting the host from autoimmune aggression, inflammation, tissue damage, and T cell transformation.
In vitro, AICD can be mimicked by stimulating T cells through ligation of the T cell receptor and addition of interleukin-2 and then by reactivation of the T cells. Various mechanisms have been described that induce AICD. In principle, chronic T cell activation leads to the upregulation of death receptors and their ligands, leading to cell death in an autocrine fashion (suicide, autonomous AICD) or by cell-cell contact (fratricide). Although several death receptors have been implicated in this mechanism, Fas and Fas ligand seem to be the most important receptor/ligand couple involved. For example, immune-privileged tissues as well as certain tumors express Fas ligand in order to avoid cell death induced by attacking CD8 cells. There are two possible ways that activated T cells can upregulate Fas ligand. First, T cell activation increases Fas ligand transcription through the induction of a number of transcription factors, including NFAT, NF-kB, and Egrl and 2.43 In addition, there are preformed Fas-ligand-containing vesicles in the cell that are released into the extracellular space and membrane upon T cell stimulation.44