THYMIC MICROENVIRONMENT
Thymocyte-Stroma Cross Talk
Developing thymocytes and thymic stroma coexist in a complicated interdependent relationship (Figure 4.3). In addition to presenting self-peptide-MHC complexes to developing thymocytes, stromal cells express costimulatory ligands and adhesion molecules that augment TCR-initiated signals and affect both positive and negative selection.
However, the functions of stromal cells during thymocyte development extend far beyond the selection phase. TECs also produce chemo- kines to direct thymocyte trafficking through the cortex and medulla. Furthermore, TECs secrete cytokines and growth factors, such as IL-7, SCF, and TSLP, that are critical for thymocyte survival and differentiation. The importance of TECs during T cell development is apparent in patients with DiGeorge’s syndrome, who have disorganized or absent thymic epithelial tissue and, consequently, exhibit profound defects in T cell production. Conversely, thymocytes play an important role in TEC survival and organization. In transgenic mice that overexpress CD3ε early in T cell development, thymocytes die prematurely, leading to thymic atrophy and disorganization of thymic epithelial tissue.72 HIV subverts these tenuous thymocyte-stroma interactions by directly killing thymocytes. The subsequent indirect effects of thymocyte depletion on the thymic microenvironment can further disrupt thymopoiesis.Disruption of the Thymic Microenvironment
HIV interacts with the thymic microenvironment in a bidirectional manner (Table 4.2). For example, the cytokine milieu established by TECs and thymocytes in the thymus promotes HIV replication. Studies have demonstrated that TECs are critical for HIV replication in thymocyte-TEC cocultures73 and that TEC-derived cytokines, such as IL-1 and TNF-α, activate NFκB, resulting in HIV transcription.74 Although TNF-α and IL-1 are primarily involved in NFκB-mediated HIV transcription, IL-7 can augment the effects of these cytokines during viral replication.75 Additionally, TEC-derived cytokines, such as IL-6 and GM-CSF, contribute to HIV replication in cultured thymocytes.76 Viral replication is also enhanced by IL-2, IL-4, and IL-7.77 Furthermore, IL-2 in conjunction with IL-4 increase expression of the co-receptors CXCR4 and CCR5 on thymocytes, whereas IL-4 and IL-7 upregulate thymocyte expression of CXCR4.78
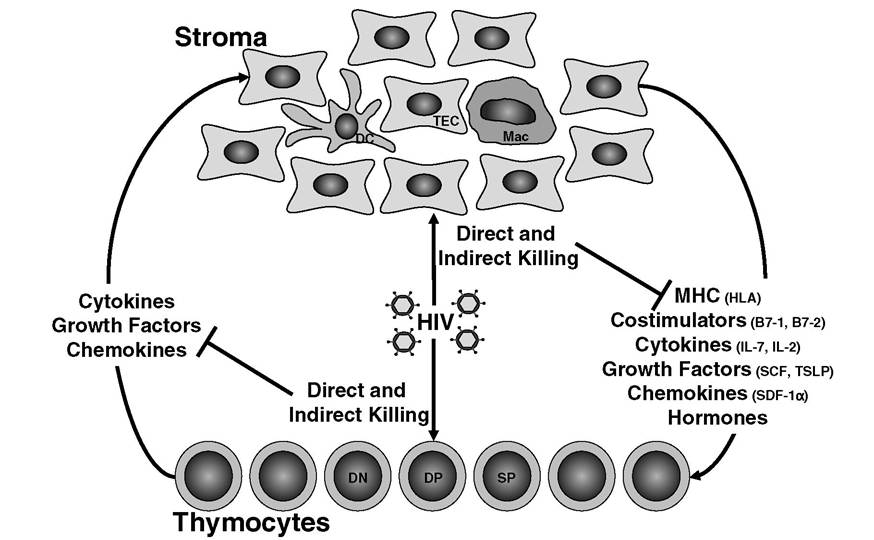
FIGURE 4.3 Thymocyte-stroma cross talk.
Thymic stromal cells and thymocytes are mutually dependent on one another for survival factors and developmental cues. HIV can directly infect and kill subsets of thymocytes and stromal cells. Consequently, thymocyte-stroma cross talk is interrupted, and HIV-mediated thymic dysfunction is amplified.TABLE 4.2
Disruption of the Thymic Microenvironment in HIV Infection
Immune Mediator Effects of/on HIV
| IL-1 | Infection increases production by macrophages in thymus; increases NFκB and viral transcription in vitro |
| IL-2 | Increases HIV replication in thymus in vitro and in vivo; upregulates CXCR4, CCR5 on thymocytes |
| IL-4 | Infection increases expression in thymocytes; increases HIV replication in thymus in vitro; upregulates CXCR4, CCR5 on thymocytes |
| IL-6 IL-7 | Infection increases expression in thymic macrophages and thymocytes; enhances viral replication Infection increases levels in serum; increases HIV replication in thymus in vitro; cofactor for IL- 1 and TNF-α in HIV transcription in thymus; upregulates CXCR4 on thymocytes |
| IL-10 GM-CSF TNF-α IFN-γ IFN-α MIP-1β Fas Bcl-2 MHCI | Infection increases expression in thymocytes Enhances viral replication Increases NFκB and viral transcription in vitro Infection increases expression in thymocytes Infection increases production by DC in thymus; upregulates MHCI on thymocytes Infection reduces expression in thymocytes Increases on thymocytes after SIV infection Decreases on thymocytes after SIV infection Decreases on thymocytes after SIV infection; increases in SCID-hu after infection |
Indirect Effects of HIV on the Thymus
The indirect effects of HIV on thymocyte development are exemplified by the observation that HIV infants born to HIV+ mothers exhibit reduced CD4 counts and decreased thymic output.79 Additional evidence suggesting that HIV indirectly disrupts thymocyte development derives from the observation that the virus directly infects far fewer thymocytes than actually die during the course of infection.80 Consistent with the latter observation, uninfected thymocytes die after contact with thymic dendritic cells induced to express FasL and TNF-α during HIV infection.81 In addition to TNF-α, infected thymic dendritic cells produce high levels of IFN-α,82 which can increase MHC class I expression on TECs and potentially influence selection of SP CD8 thymocytes.83 In addition, thymic macrophages secrete IL-1 and -6 in response to HIV infection.13 Overall, expression patterns of numerous cytokines become altered during HIV infection.
For example, SCID-hu mice infected with HIV produce excessive levels of IL-10 in the thymus.84 More detailed analysis of thymic tissue derived from infected SCID-hu mice demonstrated that thymocytes contained fewer MIP-1β transcripts and more IL-4, IFN-γ, and IL-6 transcripts than in uninfected controls.85 Similarly, thymic stromal cells from HIV+ SCID-hu mice contained more IL-6 transcripts than controls.85 In humans, IL-7 levels accumulate to abnormally high levels after HIV-mediated T cell depletion.86,87 Elevated levels of TNF-α and IL-6 have also been documented in seropositive patients, whereas IL-2 and -12 levels were reduced.88 During progression to AIDS, evidence exists for a shift in cytokine production from a Th1 profile (IL-2, -12, and IFN-γ) to a Th2 profile (IL-4, -5, -6, and -10)89,90; however, these observations were not confirmed in a majority of patients.91 Although the specific consequences of cytokine dysregulation during HIV infection of the thymus are unclear, subtle changes in the thymic microenvironment can have drastic effects on thymocyte development. In addition to cytokine dysregulation, disturbances in steroid production were reported during HIV infection. Indirect effects of HIV can decrease levels of the cortisol antagonist dehydroepiandrosterone (DHEA) and may augment steroid-induced apoptosis in thymocytes.92thymic output and t cell homeostasis
Attrition of T Cells
Thymic output as well as homeostatic expansion of the remaining T cell pool (Figure 4.4) achieves compensation for attrition of peripheral T cells. In children and young adults bearing an active thymus, the peripheral T cell pool is largely replenished by thymic output. In contrast, thymic atrophy in adults precludes thymic output as a primary mechanism for T cell replenishment, and peripheral expansion of T cells predominates in maintaining T cell numbers.
The inherent drawback of peripheral expansion is that T cells with new antigen specificities cannot be generated. Rather, existing T cells merely proliferate. Thymic output, however, enriches the T cell repertoire by producing new T cells. In order to reconstitute the peripheral T cell pool after depletion by HIV, boosting thymic output could become invaluable. However, this approach has proven difficult, as the thymus may function independently of the peripheral T cell pool. When thymic tissue was implanted into mice, the size of the endogenous thymus remained unchanged.93,94 Furthermore, cellular labeling studies performed in mice demonstrated that thymic output remains at 1 to 2% of total thymocytes regardless of the size or composition of the peripheral T cell pool.95-97 This rate of thymic output seems to be static and does not change with age, although the total numbers of T cells generated in the thymus decline as the thymus atrophies. Depending on the age of the patient, replenishment of the T cell pool after HIV infection can occur by thymic output or peripheral expansion. However, the extent of immune reconstitution may be limited due to the fact that HIV ravages both the thymus and the peripheral T cell pool.The natural course of HIV infection begins with an acute symptomatic phase lasting a few weeks that is typified by significant viremia and a rapid decrease in peripheral blood CD4 T cells.
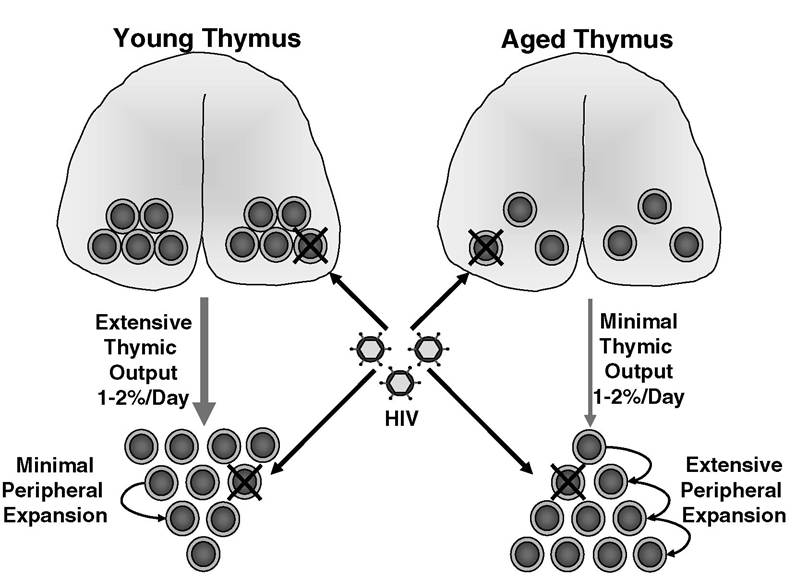
FIGURE 4.4 The impact of HIV on T cell homeostasis. HIV depletes both thymocytes and peripheral T cells. Replenishing the peripheral T cell pool can be achieved by thymic output of new T cells or peripheral expansion of existing T cells. Although young and aged thymuses produce proportionately similar numbers of T cells (1 to 2% of total thymocytes/day), output from a young thymus is greater than from an aged thymus due to the fact that a young thymus is larger and contains more total thymocytes than an aged thymus.
In a young individual, T cell homeostasis is maintained by extensive thymic output and minimal peripheral expansion. Older individuals maintain T cell homeostasis by extensive peripheral expansion and minimal thymic output.Subsequently, viremia partially subsides, and T cell counts rebound during an asymptomatic phase of chronic infection that lasts an average of 10 years. During this phase, CD4 counts gradually fall, and viremia slowly rises until the host immune system is compromised to the point at which opportunistic disease takes hold, marking the onset of AIDS. Due to technical obstacles, it has been difficult to monitor the effects of HIV on the thymus during the early stages of infection. Consequently, insight has been gained from SIV models. The initial stages of SIV infection are characterized by a reduction in peripheral blood CD4 counts correlating with suppression of thymopoiesis and increased apoptosis in the thymus.98 Approximately 2 months after infection, a rebound in thymic function occurs that is independent of viral load and lymphopenia.98 Subsequently, peripheral CD4 counts slowly dwindle throughout the course of disease,98 and at the time of progression to AIDS, thymocyte numbers are significantly reduced.99
Thymic Output
In human subjects, studies have unequivocally demonstrated that HIV inhibits thymic function. Frequently, phenotypic markers of naive vs. memory T cells are used to estimate thymic output. The presence of large numbers of peripheral T cells bearing the naive phenotype CD62Lhi CD45RA+ was interpreted to indicate that the thymus is actively producing new T cells. However, phenotypic markers are not absolute determinants of recent thymic emigrants (RTEs), as naive T cells may be long-lived, and memory CD45RO+ cells may revert back to CD45RA+ expression. A more accurate marker of RTEs is the presence of T cell receptor excision circles (TRECs).
These TRECs represent circular strands of genomic DNA generated by excision of intervening DNA sequences during rearrangement of the TCR-alpha (TCRA) locus in DP thymocytes. TRECs are remarkably stable and may be detected in RTEs by PCR. The disadvantage of using TRECs to estimate thymic output is that proliferation of RTEs results in dilution of total TRECs per cell. Consequently, TREC analysis is more appropriately used in conjunction with metabolic labeling (BrdU, deuterated glucose) or analysis of cell-cycle markers (Ki67). Consistent with this approach, TREC levels were reported to fall early in HIV infection, whereas CD4 proliferation did not rise compared with uninfected controls.100 TREC numbers are also reduced during SIV infection101 and in HIV+ children.102Peripheral Expansion
In addition to suppressing thymic output, HIV hinders peripheral T cell expansion. Metabolic labeling studies have demonstrated that the half-life of circulating T cells is drastically reduced in HIV+ patients,103 but HIV can also counteract peripheral expansion by directly infecting proliferating T cells. A delicate equilibrium exists between HIV and the T cell pool during the asymptomatic phase of chronic infection. Because HIV inhibits thymic output as well as peripheral expansion, viremia must be completely controlled or abolished in order to replenish the T cell pool. Furthermore, achieving full reconstitution of the T cell repertoire after depletion by HIV may be greatly enhanced by robust thymic activity.