THYMIC SELECTION AND T CELL MATURATION
Positive and Negative Selection
After entry into the thymus, lymphoid progenitors mature into DN thymocytes and migrate through the cortex, where they undergo selection. Developmental cues further drive DN maturation through the ISP stage and into DP thymocytes that undergo positive and negative selection at the corti- comedullary junction and in the medulla.
Here, medullary epithelial cells synthesize a myriad of tissue-specific self-antigens, the expressions of which are regulated by the transcription factor aire. These self-antigens are processed and presented to developing thymocytes by MHC on the surface of epithelial cells. Additionally, thymic antigen presenting cells such as macrophages and dendritic cells can present self-peptides to thymocytes. In this way, developing thymocytes are exposed to a broad spectrum of self-antigens during selection. Ultimately, autoreactive thymocytes that are activated by self-antigens are eliminated by negative selection, incompetent thymocytes die by neglect, and self-MHC-restricted thymocytes capable of recognizing foreign antigens mature as a result of positive selection (Figure 4.1). As such, central tolerance is achieved.Tolerance to HIV
Due to the fact that HIV antigens are presented to thymocytes in the thymus, it is relevant to question whether or not these antigens can be perceived as self-antigens and induce tolerance. Indeed, functional HIV-specific T cells are present in the periphery, suggesting that tolerance to HIV is not generated during thymic selection, that tolerance is incomplete, or that HIV-specific T cells developed before infection. Additional work will be required to determine the role of HIV infection in altering central tolerance. Recent studies addressed the possibility that HIV-encoded superantigens promote clonal deletion. One study suggested that HIV superantigens were responsible for skewed TCR Vβ profiles observed in AIDS patients; however, a subsequent report suggested that this was due to cytomegalovirus (CMV) rather than HIV superantigens.44 Also disputing the existence of HIV superantigens are data in the SCID-hu model showing that HIV infection does not preferentially deplete cells expressing specific Vβ genes.45,46 A separate possible consequence of exposure to HIV antigen in the thymus is that thymocytes become anergized or unresponsive to the virus.
Anergic T cells specific for HIV have been documented in the periphery,47 but it is not yet clear if exposure to HIV in the thymus was necessary or sufficient for anergy induction. Alternatively, HIV-specific thymocytes that encounter antigen in the thymus may develop into regulatory T cells. Upon future encounters with HIV in the periphery, these regulatory T cells could suppress immune responses to the virus. In order to clearly elucidate the pathogenesis of HIV, it will be important to further examine the potential role of HIV in generating regulatory T cells, inducing anergy, or driving clonal deletion.Thymocyte Signaling and Death Mechanisms
Central tolerance in the thymus is dependent on positive and negative selection, both of which are initiated by the TCR. Autoreactive thymocytes bearing TCRs with high affinity for self-peptide- MHC transmit strong TCR signals. In contrast, self-restricted thymocytes bearing TCRs with low affinity for self-peptide-MHC transmit weak TCR signals. Paradoxically, the strength of the TCR signal determines two opposite fates: death (negative selection) or life (positive selection). Specifically, strong signals trigger the negative selection pathway, weaker signals divert toward the positive selection pathway, and the absence of TCR signals leads to death by neglect. The TCR-proximal signals are graded and vary in intensity according to TCR avidity. This enables downstream signaling molecules to interpret upstream signals and initiate positive or negative selection as deemed necessary.
It is well documented that HIV proteins disrupt TCR signals in mature T cells48; however, little is known about how HIV influences thymocyte signaling in the context of thymic selection. Numerous mechanisms could account for HIV infection shifting TCR signals toward positive or negative selection. Specifically, CD4 engagement by HIV gp120 may activate Lck and augment TCR signals, thereby increasing negative selection.
Recent studies have demonstrated that CXCR4 can co-stimulate TCR signals and augment ERK (extracellular regulated kinase) activity,49 which regulates positive selection and maturation of DP to SP thymocytes. These studies raise additional questions as to how CXCR4 engagement by HIV gp120 may augment positive selection. Although TCR-proximal signals initiated during negative selection have been studied extensively, downstream pathways resulting in apoptosis of thymocytes largely remain a mystery. It is clear, however, that the proapoptotic Bcl-2 family members Bim, Bax, and Bak, which regulate the mitochondria- dependent apoptotic pathway, are required for negative selection50,51 (Figure 4.2). HIV-encoded Tat and gp120 may augment negative selection by facilitating Bax insertion into the mitochondrial membrane.52 HIV proteins may also hinder thymocyte survival by inhibiting the activity of anti- apoptotic Bcl-2 family members.52 Overall, the possible effects of HIV on thymocyte selection are fertile ground for investigation.TCR-initiated negative selection pathways and death by neglect are responsible for a large fraction (>95%) of apoptosis that occurs in the thymus; however, alternative apoptotic responses exist. For example, thymocytes will die after stimulation with anti-Fas antibodies or treatment with corticosteroids. Studies of TNF receptor family members (Fas, TNFR, TRAIL) in the thymus have largely focused on identifying the potential roles of these receptors in regulating negative selection. Interestingly, mice deficient in these death receptors or their ligands exhibit normal thymocyte
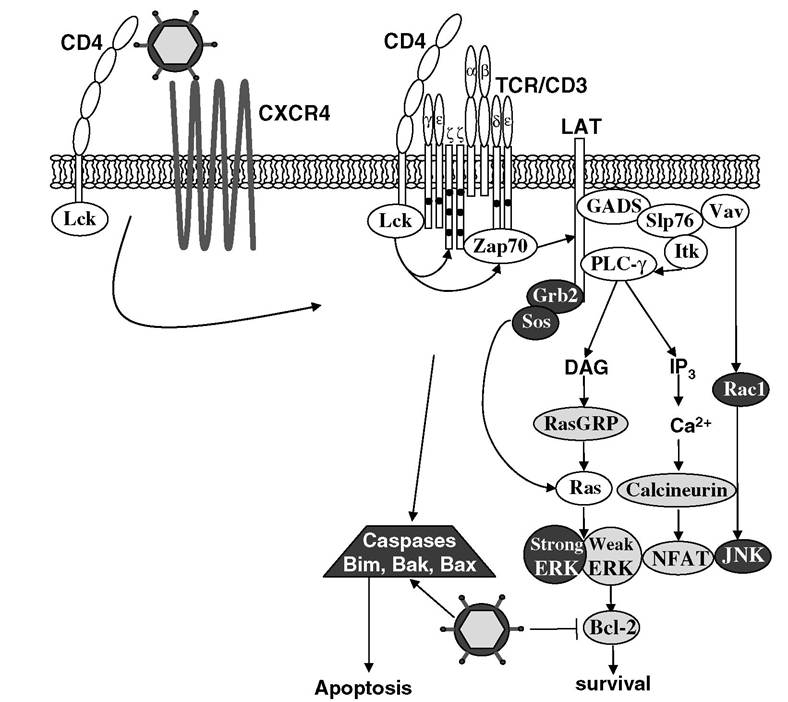
FIGURE 4.2 Thymocyte signaling. The strength of TCR signals generated in developing thymocytes dictates signaling through positive selection pathways (light gray) or negative selection pathways (dark gray). HIV may influence thymocyte survival by engaging CD4 or CXCR4 or by interfering with apoptotic regulators such as Bcl-2 family members.
For a review of TCR signaling in thymocytes, see Starr, T.K., Jameson, S.C., and Hogquist, K.A., Annu. Rev. Immunol., 21, 139, 2003.development.53-54’55 In addition, mice defective in signaling molecules downstream of death receptors (transgenic mice expressing dominant negative Fas-associated protein with death domain [FADD],56 caspase-9 knockouts,57 and Apaf-1 knockouts58) do not exhibit defects in negative selection. Together, these studies indicate that death receptors do not mediate TCR-initiated negative selection. However, HIV infection could potentially enhance thymocyte susceptibility to death receptor-initiated apoptosis. In the SIV model, infection increases Fas expression and reduces levels of Bcl-2 in thymocytes.59 HIV proteins may also augment apoptotic signals in thymocytes by employing mechanisms similar to those used in peripheral T cells (see Chapter 5 in this text), but this is yet to be formally demonstrated.
Direct evidence for HIV-encoded proteins disrupting survival signals in thymocytes was obtained from transgenic mice. A mouse expressing HIV provirus lacking Gag, Pol, and Env exhibited thymocyte depletion despite the fact that the provirus was replication incompetent.60 Evidence that Nef may play a pivotal role in HIV-mediated thymocyte toxicity is supported by the observation that excessive thymocyte depletion occurs in Nef transgenic mice.61 Furthermore, the thymocytes that survived were hyperresponsive to activation by anti-CD3 antibodies.62 Subsequent analysis of Nef transgenic mice revealed that DP thymocytes express low levels of CD4.63 Interestingly, Nef toxicity in thymocytes correlates with reduced CD4 expression.64 In the SCID-hu model, HIV bearing a Nef mutation that does not affect viral replication failed to decrease CD4 expression and was less toxic to thymocytes than was wild-type Nef.65 Several additional studies confirmed that HIV Nef mutants are less pathogenic than wild-type viruses in the SCID-hu thymus.66,67 In human thymocytes, expression of Nef impairs thymopoiesis in vitro68 and leads to excessive thymocyte proliferation.69 Ultimately, Nef may mediate its toxic effects by nonspecifically activating thymocytes and by downregulating CD4, but Nef has also been shown to kill cells by inhibiting expression of the antiapoptotic molecules Bcl-2 and Bcl-xL.52 Similarly, Tat can inhibit Bcl-2 expression, but it activates the proapoptotic factor Bax as well.52 Although Tat expression in thymic stromal cells inhibits thymocyte maturation in vitro,10 transgenic mice expressing Tat under the control of a CD2 promoter develop normally.71 Similar to Tat, gp120 can activate Bax and inhibit Bcl-2 expression.52
From a broad perspective, multiple stimuli can initiate thymocyte death.
In addition to negative selection, thymocytes die by neglect, death receptor engagement, exposure to corticosteroids, and cytokine deprivation. HIV disrupts the thymic microenvironment at the cellular and molecular levels, resulting in dysregulation of developmental cues and survival signals required by thymocytes.