VIRAL PROTEIN R (VPR): STRUCTURE AND FUNCTION IN THE VIRAL LIFE CYCLE
Genomic Structure of Vpr in HIV-1
Vpr constitutes one of the four HIV accessory proteins that include Nef, Vpu, and Vif. Its expression late in the viral life cycle requires the specific interaction with the C-terminal domain of p55 Gag19, and approximately 100 copies are incorporated into each virion.1 Vpr is highly conserved among HIV-1, HIV-2, and simian immunodeficiency virus (SIV).2 The full structure of Vpr has not been determined given its tendency to aggregate and multimerize in solution.
As an alternative approach,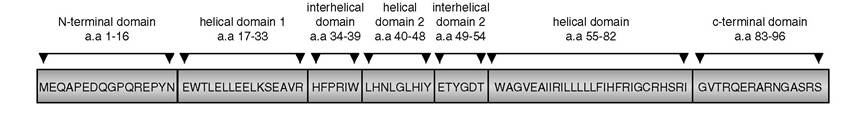
FIGURE 7.1 Amino acid structure and domains of HIV-1 Vpr.
synthetic peptides were used to analyze several Vpr domains by circular dichorism and homonuclear and heteronuclear nuclear magnetic resonance (NMR) methods.3-6 Its structure is characterized by several tandem repeats of γ-turns and α-helices. Mutagenesis studies indicate that the C-terminal domain is responsible for cell-cycle arrest and protein stability,7’8 and it contains several acidic motifs that play a role in its cytotoxicity.9-11 The N-terminal domain is required for nuclear localization, virion packaging, and binding of a number of viral (nucleocapside protein NCp7, p17) and cellular proteins (RIP1, Sp1).12,13 Based on this information, Vpr domains mediate distinct functions and may explain the pleiotropic effects of its action during the viral life cycle.
Amino Acid Sequence
Vpr is a 14 kDa protein composed of 96 amino acids divided into several domains. The key domains are shown in Figure 7.1. The N-terminal (amino acids 1 to 16) forms the first γ-turn. Sequence variations in this region among HIV-1 clades were noted.14 This is followed by a helical domain (amino acids 17 to 33), which by specific site-directed mutagenesis was shown to be important for subcellular localization, virion incorporation, stability, and oligomerization.15-19 In particular, an amino acid substitution at residue 33 was isolated in naturally occurring viruses that dramatically affects viral subcellular localization and virion incorporation.20 A second γ-turn (amino acids 34 to 39) is found between the first and second helical domains.
Although some natural mutations were isolated, further characterization is needed to determine their functional consequences on Vpr activity.21 The second helical domain forms a highly hydrophobic region that significantly abrogates virion incorporation when mutated.16 The second interhelical domain spans the third γ-turn comprising amino acids 49 to 54. Although naturally occurring polymorphisms are found in this region, its contribution to Vpr function is not known. The last helical domain (amino acids 55 to 82) was implicated in both virion incorporation and subcellular localization. This region contains an arginine- rich motif that, when expressed in yeast or mammalian cells, exhibits potent apoptotic-inducing activity.7,20,22-24 The C-terminal region of Vpr participates in cell-cycle arrest with four serine residues that are targets for phosphorylation.25Lessons from Yeast
A large number of cellular functions are highly conserved in both yeast and mammalian cells. This has led to a number of focused studies on Vpr function using fission yeast Schizosaccharomyces pombe and budding yeast Saccharomyces cerevisiae (reviewed in Zhao and Elder2). In yeast, progression through G2 requires phosphorylation of Cdc2 on tyrosine 15 by the Wee1 and Mik1 kinases.26,27 Genetic studies suggest that Vpr induces G2 arrest by inhibiting phosphorylation of Cdc2 kinase.26,28-31 This effect is distinct from physiological or pharmacological forms of DNA damage or replication checkpoint pathways. DNA damage in yeast activates the Cdc2 pathway, whereas the checkpoint pathway involves Rad1, Rad3, Rad9, and Rad17, which lead to phosphorylation of Chk1 kinase. Chk1 directly phosphorylates Cdc25 phosphatase,31 allowing the transport of Cdc25 out of the nucleus, rendering it inactive.26 Vpr induces G2 arrest in chk1∕cds1 double-yeast mutants, supporting the hypothesis that the mechanism of Vpr action is distinct from DNA damage or checkpoint regulation.
Interestingly, another protein that regulates cell-cycle progression, PP2A, plays a major role in G2 arrest.28 PP2A interacts with both Weel and Cdc25 in S. pombe, and its overexpression results in G2 arrest. The PP2A inhibitor okadaic acid suppresses Vpr-induced G2 arrest, and deletion of one of the catalytic PP2A subunits in S. pombe has a similar negative effect.32The cytotoxic domain of Vpr (HFRIGCRHSRIG) was originally described in S. cerevisiae and S. pombe systems.23 Using synthetic peptides, the authors showed that the cytotoxic domain induced cell-cycle arrest and loss of mitochondria membrane potential. Only recently was a similar comprehensive study performed in mammalian cells. In fact, the data on cell-cycle targets recapitulated what was found in the yeast system. Because yeast lacks the core apoptotic components of the mitochondria-induced cell death pathway, the more recent work provided important data regarding the aspects of Vpr-induced T cell death.