Placentation and normal placenta development
The development of the placenta starts from implantation, when the embryonic pole of the blastocyst enters into contact with the maternal uterine epithelium. Human placentation is almost unique amongst mammals in that it is highly invasive and the gestational sac embeds itself completely within the uterine endometrium and superficial myometrium (5, 8).
Human placentation is also characterized by remodelling of the spiral arteries, during which the vessels lose their elastic lamina and smooth muscle coat, and consequently their responsiveness to circulating vasoactive compounds. At term, the spiral arteries that, in the non-pregnant state, transport just a few millilitres of blood per minute need to carry up to 600 mL per minute.Implantation, decidualization, and anchoring
The formative stages of human placental development are largely unknown. The earliest events have never been studied in vivo for obvious ethical reasons (9). Implantation relies on the interaction of the trophectoderm cells forming the wall of the blastocyst with the cells of the uterine epithelium. On establishing contact, some of the trophectoderm cells undergo proliferation and fusion to form the multinucleated syncytiotrophoblast, whereas others remain as a deeper, progenitor population, the cytotrophoblast cells (10).
During the secretory phase of the menstrual cycle, the endometrium transforms into a well-vascularized receptive tissue, which is characterized by the proliferation and differentiation of the stromal cells into decidual cells, the infiltration of maternal immune cells, and vascular remodelling of the endometrial vessels (11, 12). Decidualized stromal cells are derived from the fibroblast-like cells within the endometrium. The uterine glands undergo characteristic hypersecretory morphological changes, the so-called Arias-Stella reaction under the influence of progesterone (13).
Their cytoplasm contains abundant organelles and large accumulations of glycogen, an important source of nutrients for the early embryo. Their secretions are also rich in growth factors such as leukaemia inhibitory factor, vascular endothelial growth factor, epidermal growth factor, and transforming growth factor beta, which all play a role in placentation (14).Soon after implantation, projections of syncytiotrophoblast (primitive villi) penetrate between the epithelial cells of the uterine decidua, while at the same time the endometrial stromal cells grow over and encapsulate the whole gestational sac. Strands of mononuclear cytotrophoblast start to proliferate at the fetal side of the implanted blastocyst wall (9, 10). The resulting cytotrophoblastic columns push themselves into the primitive syncytiotrophoblastic mass to form the secondary placental villi. The most distal cytotrophoblast cells break through the syncytium and spread laterally to form the cytotrophoblastic shell separating the placenta from the decidua. Cells on the outer surface of the shell differentiate into non-proliferative, cytotrophoblast cells that invade the decidual stroma, collectively called extravillous trophoblast (EVT). They differentiate primarily into interstitial and endovascular subpopulations that migrate through the decidual stroma and down the lumens of the spiral arteries respectively. The interstitial EVT invade the uterine wall as far as the inner third of the uterine myometrium, where they fuse to form multinucleated trophoblast giant cells (15). This area is known as the junctional zone.
Development of the uteroplacental circulation
The left and right uterine arteries are the main blood supply to the uterus. They ascend along the lateral aspects within the broad ligament and terminate by anastomosing with the corresponding ovarian artery. The uterine arteries give rise to arcuate arteries that pass medially and penetrate the myometrium and divide into anterior and posterior branches that run circumferentially between the outer and middle thirds of the myometrium, and anastomose freely with their counterparts from the opposite side in the midline (Figure 9.2).
The arcuate arteries give rise to the radial arteries that are directed towards the lumen of the uterus. As they reach the junctional zone, each radial artery gives off lateral branches, the basal arteries that supply the myometrium and the deeper basalis parts of the endometrium, and continues as a spiral artery (16).The spiral arteries are highly coiled within the decidua basalis and the deeper parts of the functionalis. As they approach the uterine cavity they rapidly narrow, and divide into several smaller branches that follow a straighter course before terminating in a capillary plexus just beneath the uterine epithelium. Each spiral artery also gives off small branches supplying the capillary plexus surrounding the The EVT cells penetrate the junctional zone via the action of their proteases on the intercellular ground substance, affecting its mechanical and electrophysiological properties, and changing the structure of the walls of the spiral arteries. EVT cells can first be found both within and around the spiral arteries in the central area of the placenta. They gradually extend laterally, reaching the periphery of the placenta around mid gestation. Depth-wise, the changes are maximal within the central region of the placental bed and the extent of invasion is progressively shallower towards the periphery.
The remodelling of the spiral arteries is characterized by the progressive loss of myocytes from their media and of their internal elastic laminae, which are replaced by fibrinoid material (10, 17). Consequently, these vessels lose their responsiveness to circulating
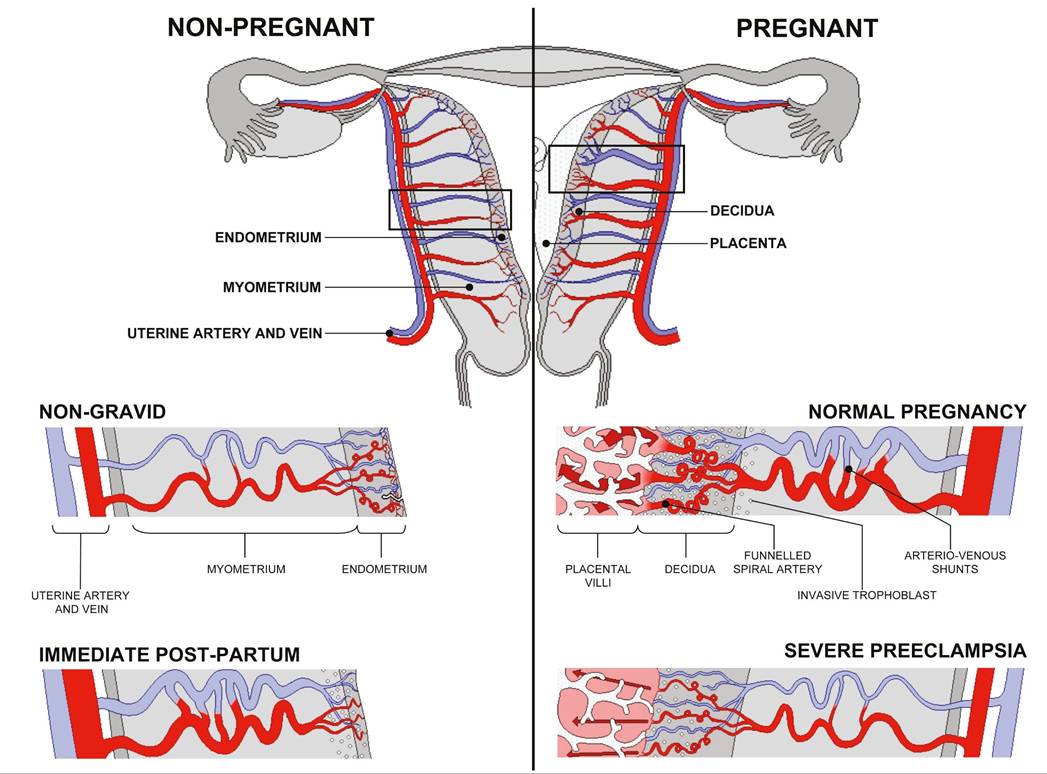
Figure 9.2 Diagrammatic representation of uterine and placental vasculature (red shading, arterial; blue shading, venous) in the non-pregnant, pregnant, and immediate postpartum state. Normal pregnancy is characterized by the formation of large arteriovenous shunts that persist in the immediate postpartum period.
By contrast, pregnancies complicated by severe pre-eclampsia are characterized by minimal arteriovenous shunts and thus narrower uterine arteries. Extravillous cytotrophoblast invasion in normal pregnancy (diamonds) extends beyond the decidua into the inner myometrium resulting in the formation of funnels at the discharging tips of the spiral arteries. Contrast with severe pre-eclampsia.Reproduced from Burton GJ, Woods AW, Jauniaux E, Kingdom JC. Rheological and physiological consequences of conversion of the maternal spiral arteries for uteroplacental blood flow during human pregnancy. Placenta. 2009;30:473-82 (https://creativecommons.org/licenses/by/3.0/).
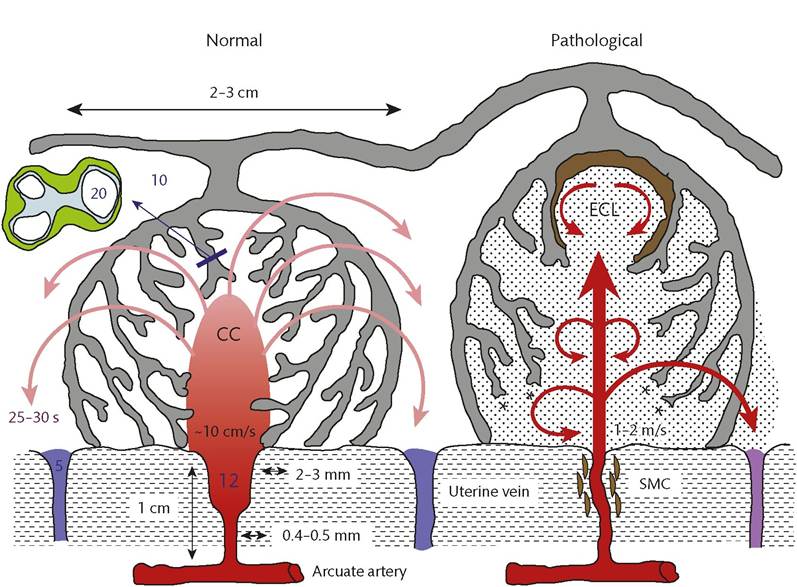
Figure 9.3 Diagrammatic representation (not to scale) of the effects of spiral artery conversion on the inflow of maternal blood into the intervillous space in normal pregnancy and in pregnancies complicated by insufficient transformation of the spiral arteries. The retention of smooth muscle cells (SMCs) around the spiral artery will increase the risk of spontaneous vasoconstriction and ischaemia-reperfusion injury CC, central cavity; ECL, echogenic cystic lesions.
Reproduced from Burton GJ, Woods AW, Jauniaux E, Kingdom JC. Rheological and physiological consequences of conversion of the maternal spiral arteries for uteroplacental blood flow during human pregnancy. Placenta. 2009;30:473-82 (https://creativecommons.org/licenses/by/3.0/).
vasoactive compounds and become a low-resistance vascular network by dilatation. This transformation, termed ‘physiological changes', results in the metamorphosis of small-calibre spiral vessels into flaccid distended arteries with a five- to tenfold dilation at the vessel mouth (Figures 9.3 and 9.4). Around 30-50 spiral arteries are transformed during the first trimester. In normal pregnancies, the transformation of spiral arteries into uteroplacental arteries is
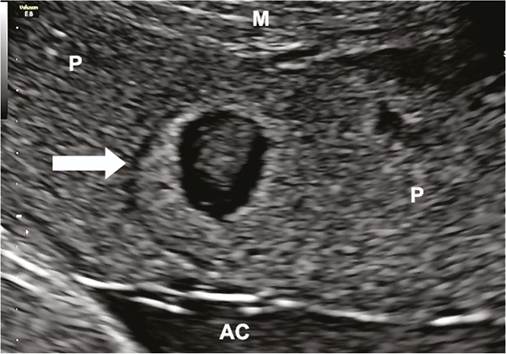
Figure 9.4 Ultrasound view of an echogenic cystic lesion (arrow) in a pregnancy at 36 weeks complicated by late-onset pre-eclampsia.
AC, amniotic cavity; M, myometrium; P placenta.described as completed around mid gestation (17). These major anatomical changes are associated with a fall in resistance to blood flow in the uterine arterial circulation as observed by Doppler ultrasonography, and a reduction in the velocity and the pressure of the maternal blood entering the placenta (17-19). However, there is a gradient in the infiltration of the EVT along the spiral artery and even in normal pregnancies, not all spiral arteries are completely transformed (17).
The early or primitive placenta
During the fifth week of pregnancy (third week after conception) fetal capillaries develop in the mesenchymal core of the secondary villi, transforming them into tertiary villi (20). By 6 weeks, the villous vasculature is connected with the primitive heart and the vascular plexus of the yolk sac via the vessels of the connecting stalk. The fetoplacental circulation is established from around 8 weeks of gestation but placental capillary formation will only be completed by mid gestation (20). These anatomical changes can explain the major changes in blood flow velocity waveforms obtained from the umbilical artery during that period (18). Lateral projections from the main stem villus branch repeatedly to form intermediate, and finally terminal, villi. Collectively, these are often referred to as floating villi as they are not attached to the decidua.
From the third week until about the second month of pregnancy, the entire chorion of the primitive placenta is covered with villus stems (20). Placental villi form initially over the entire surface of the chorionic sac, but starting towards the end of the first trimester the
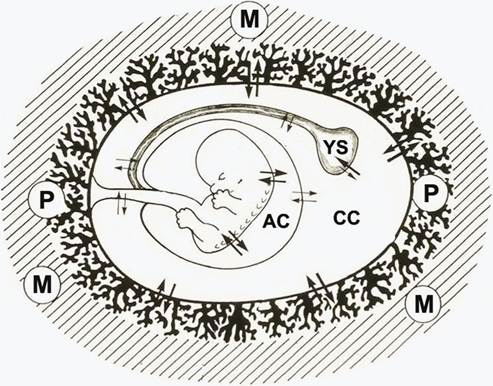
Figure 9.5 Diagram of a normal gestational sac at 8 weeks showing the outside to inside by the placental villi (P), the chorionic cavity (CC) containing the secondary yolk sac (YS), and the amniotic cavity (AC) containing the fetus.
M, myometrium.villi over the superficial pole regress, leaving the villi of definitive placenta to continue to grow. At 8 weeks, the typical normal gestational sac is composed from outside to inside by the placental villi, the extraembryonic coelom or chorionic cavity, which contains the secondary yolk sac, and the amniotic cavity, which contains the developing fetus (Figure 9.5). The secondary yolk sac is the first structure that can be detected ultrasonographically within the chorionic cavity. Its diameter increases slightly between 6 and 10 weeks of gestation, reaching a maximum of 6- 7 mm after which it decreases (21).
Anatomically, in vivo and in vitro studies have shown that the EVT cells not only invade the uterine tissues and wall of the spiral arteries but also form a continuous cytotrophoblastic shell at the interface with the decidua. This shell and the trophoblastic plugs that extend from it into the mouths of the spiral arteries act like a labyrinthine interface that filters maternal blood, permitting a slow seepage of plasma, but no true blood flow, into the intervillous space (22, 23). This has led to the concept that human placentation is in fact not truly haemochorial in early pregnancy (Figure 9.6). During the period when there is no true maternal blood circulation entering the placenta, the uterine glands deliver nutrient-r ich secretions through the shell directly into the intervillous space. These secretions are a heterogeneous mix of maternal proteins, carbohydrates, including glycogen, and lipid droplets, and are phagocytosed by the syncytiotrophoblast indicating a histotrophic nutrition process (24). They also contain a variety of growth factors, and so create a highly stimulatory microenvironment within the developing placenta.
During the ‘plugging phase' of the spiral arteries, there is physiological hypoxia inside the gestational sac, which is essential for normal placental and fetal development to occur (25, 26). This paradox reflects the fact that organogenesis is characterized by cell lineage differentiation and morphogenetic events that are critical for the development of a healthy fetus. Disruption of signalling pathways or oxidative damage to the fetal DNA at this stage will have profound consequences, and so maintaining metabolism at a low level must be advantageous (27). This concept is consistent with our
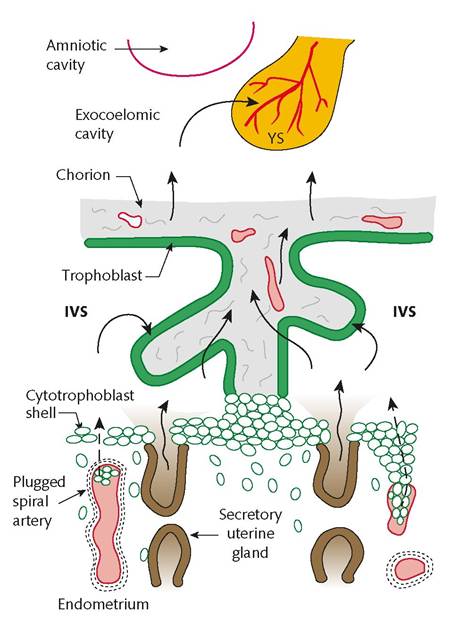
Figure 9.6 Diagram showing the histotrophic nutrition pathways in the first-trimester gestational sac. Note that there is no true maternal blood circulation entering the placenta and that the uterine glands deliver their secretions through the trophoblastic shell directly into the intervillous space (IVS). Nutrients are then transferred through the trophoblast of the placental villi into the exocoelomic cavity and into the secondary yolk sac (YS).
finding that during the first trimester placental metabolism is heavily dependent on phylogenetically ancient pathways that are less reliant on oxidative phosphorylation and so reduce the risk of generation of dangerous oxygen free radicals (27). The plugging of the spiral arteries also creates a uteroplacental oxygen gradient which exerts a regulatory effect on placental tissue development and function. In particular, it influences cytotrophoblast proliferation and differentiation along the invasive pathway, villous vasculogenesis, and the formation of the chorion laeve or free placental membranes (26, 27).
The definitive placenta
At the end ofthe first trimester, the EVT plugs ofthe cytotrophoblastic shell are progressively dislocated, allowing maternal blood to flow more freely and continuously within the intervillous space (22, 23). This transforms the placenta into a haemochorial organ. During this transitional phase, which takes place between 10 and 14 weeks of gestation, two-thirds of the primitive placenta disappears and the chorionic cavity is obliterated by the growth of the amniotic sac (26). These events bring the maternal erythrocytes closer to the fetal tissues, facilitating nutrient and gaseous exchange between the maternal and fetal circulations.
The definitive placenta is composed of a chorionic plate on its fetal aspect and a basal plate on the maternal aspect, and an intervening intervillous space containing villus stems with branches in contact with maternal blood. Towards the end of the first trimester, the villi begin to differentiate into their principal types. The connections to the chorionic plate become remodelled to form stem villi, which represent the supporting framework of each villus tree (20). These progressively develop a compact fibrous stroma, and contain branches of the chorionic arteries and accompanying veins. After several generations of branching, stem villi give rise to intermediate immature and mature villi. Mature intermediate villi provide a distributing framework, and terminal villi arise at intervals from their surface. The terminal villi are the main functional units of the villus tree. They are highly vascularized, but by capillaries alone, and are highly adapted for diffusional exchange. This differentiation of the villi coincides temporally with the development of the lobular architecture, and the two processes are most likely interlinked. Lobules can be first identified during the early second trimester, following onset of the maternal circulation when it is thought haemodynamic forces may shape the villus tree (17). Each lobule represents an individual maternofetal exchange unit.
At term, the human placenta is usually a discoid organ, 15-20 cm in diameter, approximately 3 cm thick at the centre, and weighing on average 450 g (20). Macroscopically, the organ consists of two surfaces or plates; the chorionic plate to which the umbilical cord is attached, and the basal plate that is attached the uterine wall. The placenta is incompletely divided into between 10 and 40 lobes by the presence of septa created by invaginations of the basal plate. The septa are thought to arise from differential resistance of the maternal tissues to trophoblast invasion, and may help to compartmentalize and hence direct maternal blood flow through the intervillous space (17). The maternal blood percolates through this network of channels, exchanging gases and nutrients with the fetal blood circulating within the villi, before draining through the basal plate into openings of the uterine veins.