Ecology and Evolution
While philosophers have paid attention to systematics since the 1960s, they have largely neglected ecology, despite its historical and conceptual richness. This is beginning to change, as works by Cooper (2003), Ginzberg and Colyvan (2004) and Sarkar (2005) attest.
One developing area of interest concerns the intersection of ecology with evolution. Two questions can be distinguished:• What are the relationships between evolutionary and ecological processes?
• What are the relationships between evolutionary and ecological theory?
In this section, we will explore the relationships between ecology and evolution with regard to both of these questions. The way we answer the former will inform how we approach the latter. We begin by sketching out a common and widely held view about how evolution and ecology are related, then move on to some important concerns about this view and what they might mean for theory in evolution and ecology.
4.1 A two-scale approach
One common way of thinking about how these two disciplines and their systems interact comes from a metaphor first articulated by the eminent ecologist and limnologist G.E. Hutchinson in 1957, who claimed that the “ecological play occurred in the evolutionary theatre.” By this he meant that ecological processes occur at rates and scales “below that” of evolutionary processes. To see the contrast, consider two common ecological phenomena: competition and speciation. We say that two species compete when each reduces the rate of growth of the other.[102] More specifically, biologists identify two types of interspecific competition. Interference competition occurs when individuals of a species fight with individuals of a different species. By contrast, exploitative competition occurs when conspecifics use a common limited resource. Ever since Darwin (1859), biologists of many disciplines have studied competition as it takes place, on scales from a few square meters between two organisms to much larger areas with many more organisms at the ecosystem level.
Now consider speciation, the set of processes by which new species arise. Speciation generally takes thousands of generations to occur. For example, in allopatric speciation, a formerly contiguous population of organisms is geographically divided into two or more populations. Over many generations these populations will begin to vary differentially, often because of differences in the selection process as it acts on each population. If and when these two populations meet, they may well find themselves repro- ductively isolated from each other; that is, members of the two populations may not mate, or they may mate but fail to produce fertile offspring, for any number of reasons.[103]
In the present context what is salient about these two processes - interspecific competition and speciation - is that they are generally understood to occur on different spatial and temporal scales. Competition can occur in a few square meters (look out your window and notice the small birds and mammals foraging away or fighting and you probably see instances of interspecific competition), and within one generation, but the speciation process is generally “splayed out” in space and time.[104]
The philosopher Elliott Sober gave a philosophical account of evolutionary theory that fits Hutchinson’s two-scale approach rather well. Sober (1984b) articulated and defended the view that evolutionary theory is a “theory of forces.”[105] This view is committed to several claims. First, single evolutionary processes like natural selection, mutation, migration, and drift are forces, or better yet, causes of changes in gene frequencies. These causes are customarily modeled as singleton laws. This is a way of describing how natural selection operates when no other cause is at work.[106] For example, consider a single locus on a chromosome with two alleles, A and a. There are three possible genotypes AA, Aa, and aa and we can associate with each a fitness wAA, wAa, and waa respectively.
Likewise, let us assume that the frequency of A is p and the frequency of a is q (wherep + q = 1). If we further assume that the only force at work is natural selection, then we have the following dynamical equation for a change in p: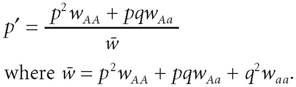
Second, these component forces can be combined or “vector summed” together into net forces. These are called laws of composition. For example, we can include mutation in our model. Let /d be the fraction of A alleles that mutate into a alleles. Thus, (1 - d) is the fraction of A alleles that do not mutate. Our new composition law for natural selection and mutation is:
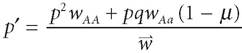
According to Sober, the models of evolutionary genetics are consequence laws because they predict how gene frequencies will change given the laws and the relevant inputs into the equations. However, the inputs - the values of the parameters (genotypic fitnesses and mutation rate) and variables (allele frequencies) - are specified exogenously. Sober claims that it is the business of ecology to provide those inputs or explanations of their values. Hence, in ecology, we find source laws. On this view, ecology explains how organisms come to differ in their fitnesses, while evolution determines what these fitness differences mean over the long term.
Here is another example of source laws in ecology from what is called “life history theory.” Let us say that an organism is semelparous if it breeds only once and an organism is iteroparous if it breeds more than once. An instance of the difference between these two reproductive strategies might be familiar from the distinction between annual and perennial plants. Given that these strategies are quite different, we might want to know whether it is better from an evolutionary perspective to be an annual or a perennial plant.[107] Consider an annual species and a perennial species and let Ba and Bp be the number of seeds that germinate in the next year respectively.
Likewise, let Na and Np be the number of annual and perennial plants respectively. Let the survival rate of the perennial species be s and the mortality rate for seeds and seedlings be s,. For the annuals and perennials, respectively, the equations that describe population growth with time are: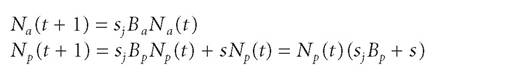
Suppose the growth rates of annuals and perennials are equal, or[108]
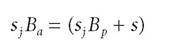
In effect, we are assuming that being an annual or perennial makes no difference. Rearranging, we have:
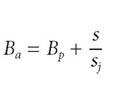
However, if we compare the survival rate of mature adults to that of seedlings, the ratio s/s, is likely to be very large (i.e., s/s, >> 1). Hence, for
the “annual strategy” to be better evolutionarily than the “perennial strategy”, Ba >> Bp. That is, the annuals must produce many, many more seeds than the perennials. Thus, under these circumstances, being a perennial will likely be favored over being an annual.
4.2 Messy interactions: concerns about the two-scale approach
While these models are useful and have been important in the development of evolutionary and ecological thinking, the approach articulated by Hutchinson and elaborated implicitly by Sober is not so tidy empirically. Biologists have found increasing evidence that ecological and evolutionary processes are commensurate in scale. Consider evolutionary processes first. It has been demonstrated that changes in beak size in Darwin's finches can occur remarkably quickly due to changes in rainfall (Grant 1986). Rapid adaptation in insects to insecticides like DDT has also been well documented. By the same token, ecological processes can occur on very long time scales. Consider a mature forest where there has been a forest fire or other disturbance.
Some species will go locally extinct and others will migrate into the area. There will be competition for nutrients and light and it may take centuries for the different frequencies of tree species to settle down into something like the previous non-disturbed distribution.Even a view on which ecological and evolutionary processes occur on the same spatiotemporal scales may still be too simplistic because it is possible for ecological and evolutionary processes to interact or causally affect one another. Niche construction is one example of this kind of interaction. According to niche construction theory, organisms can radically alter their environments in a variety of ways.[109] They can alter how matter and energy move through ecosystems (these organisms are “ecosystem engineers”). For example, there are several species of fungus-growing ants of the tribe Attini, the best known of which are leaf-cutter ants in the genera Atta and Acromyrmex. Species of these genera cut and move vegetation into nests in order to grow fungus. This provides them with a source of food, and these colonies can reach enormous sizes.
Other organisms can radically change their selective environments. The common cuckoo, Cuculus canorus, for example, places its newborns into nests of other species. These newborns then parasitize the brood in the new nest. This behavior has led to important traits such as fast incubation times and the instinct in hatchlings to push “native” eggs from the nest. The cuckoo chicks even exhibit calls that mimic the rate and sound of an entire brood, thereby causing the “mother” to feed only them. As another example, consider the evolution of photosynthesis in bacteria. Before the appearance of these organisms, the earth's atmosphere contained much less oxygen. Oxygen increase as a result of bacteria photosynthesis radically affected biological evolution thereafter.
If these examples are representative of evolutionary and ecological dynamics, then we cannot separate the “evolutionary play” from the “ecological theatre” in the customary way.
These concerns about process bring us to the question about theory. Traditionally, theorists constructed models as if Hutchinson's slogan was correct. Ecologists describe their populations of specific species in terms of their demographic properties such as birth and death rates, per capita rates of growth, interaction coefficients, and species densities. In these models, the environment of a population or community is allowed to vary with time. Theoretical ecologists took Hutchinson's slogan to heart and ignored what evolutionary geneticists understand as evolution - change in the frequencies of genes. These geneticists did something very similar as well: they wrote models that predicted the frequency of genotypes as a function of evolutionary “forces” such as natural selection, inbreeding, mutation, migration, genetic drift, etc. Moreover, they have tended to focus on single species and to assume that the environment in which a species finds itself is relatively unchanging. If Hutchinson's slogan is misleading of at least many important biological systems, then we must somehow deal with ecological and evolutionary processes occurring in multispecies assemblages in varying environments.As an illustration of how to model such assemblages, consider a very simple mathematical model of Richard Lewontin's (1983). According to Lewontin, traditional evolutionary theory can be understood in terms of the following pair of differential equations:
id="Picutre 52" class="lazyload" data-src="/files/uch_group76/uch_pgroup320/uch_uch7391/image/image045.jpg">
These equations say that organisms change over time through some functional relationship between themselves and their external environment. However, the environment changes only as some function of itself. In Lewontin's terms, the environment provides “problems” for organisms to “solve”; however, those problems remain unaffected by the organisms. Lewontin argues that this model is wholly inadequate for some of the same reasons as those mentioned above. As an alternative, he offers the following model:
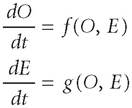
Organisms and environments codetermine each other if processes like niche construction occur frequently. Thus, the phenomena of adaptation - the fit between organism and environment - can be accomplished in more than one way. Through heritable variations in fitness, organisms can “fit” their environments better than alternative types of organisms. However, organisms can also alter their environments, thereby reversing the direction of fit, and forcing it to fit them.
In the end, the processes of evolution and ecology are not necessarily separate either in their spatiotemporal scale or interactively. This suggests that current evolutionary and ecological theory will have to be revised, possibly in radical ways. How radical these revisions should be remains to be seen.
5.