Systematics
Systematics can be considered to have two major goals: (1) to discover and describe species and (2) to determine the phylogenetic relationships of these species. (Wiens 2007, 875)
Evolutionary biologists study both the process and pattern of evolution.
Systematists primarily focus on the latter. Studying the pattern of evolution cannot be done without staking out positions that are inherently philosophical in nature. This section is a brief survey of three core issues that must be addressed in some way or another for systematics work to move forward, and are the kinds of questions that represent the overlapping interests of philosophers and biologists.In order to discover species, systematists must have some idea of what it is to be a species. This turns out to be not simply a biological question, but a deeply philosophical one as well. Furthermore, providing a coherent and useful description means conveying the relevant criteria that suggest that something is, indeed, a species. Determining the phylogenetic, or genealogical, relationships between groups of species requires making an inference about the distant past that is not directly observable. Some justification of inferences that places these claims within the proper theoretical (evolutionary) framework is needed if they are to be scientifically plausible. These challenges are both metaphysical and epistemological in nature. Let us look at each a bit more carefully.
3.1 The species category
For a long time, species were taken to be exemplars of natural kinds (i.e., sets; see below) by philosophers. Now, however, things are not so simple. What this means, and why this matters for both philosophy and biology, is a classic example of a philosophy of biology problem.
Sets are abstract entities defined by their membership. Traditionally understood, sets may be characterized by necessary and sufficient properties, possession of which determines membership in a set.
In other words, something is a member of a set by virtue of instantiating the defining properties of that set. If species are sets, then belonging to a species means satisfying conditions for membership in that set, that is, possessing the defining characters of that species. It also means, at least in a traditional sense, that species are abstract entities, and good candidates for being natural kinds.This characterization of species is at odds with Darwinian thinking, because as Mayr (1959) and Sober (1980) have pointed out, an important feature of Darwinian theory is that populations vary in ways that affect fitness (see §5 below). That is, evolutionary theory requires dynamics that the natural-kind view denies. In response, Michael Ghiselin (1966, 1974) and David Hull (1976, 1978) have forcefully argued that particular species are best understood as individuals, as opposed to sets or classes. The individuality thesis is straightforward: the species category (or taxon) is a class, with individual species as members.[99] A species is an individual, as opposed to a set. This means that species are best understood as having parts, as opposed to members, and that species are historical entities, which is to say that they exist in space and time, rather than abstractly. Individual organisms are parts of particular species rather than members of them.
Belonging to a species, then, means being a part of some historical entity, albeit one that is more scattered than other, perhaps more familiar, individuals like organisms. Notice that “being an individual” is not coextensive with “being an organism” though the two notions are often used interchangeably (Hull 1978, Wilson and Sober 1989, Hamilton, Haber, and Smith 2009). Certainly being an organism entails being an individual, but the reverse is not true.
The timing of the individuality thesis was good. Biologists were generally receptive to it, as it accorded well with shifts in biological practice, namely a general move towards phylogenetic thinking occasioned by the work of German entomologist and theorist Willi Hennig (1966).
Taxonomists placed less emphasis on traits and more emphasis on history (or other extrinsic properties) as the theoretically relevant feature of what it meant to be a particular taxon, meaning that taxa are defined by ancestry, not possession of any particular features. The individuality thesis provides a conceptual basis for this position - rather than looking for some essential property, taxonomists instead can specify what it means to be a part of a species in the relevant way (more on this below).Philosophers have been more resistant to the individuality thesis than most biologists, though generally receptive. Critics of the individuality thesis have complained, among other things, of a conflation of thinking of species as sets with the thesis that species are natural kinds, or that either entails some sort of essentialism (Boyd 1999, Griffiths 1999, Wilson 1999b, Winsor 2006). Other philosophers have argued that thinking of species as sets is not at odds with evolutionary theory, as proponents of the individuality thesis have suggested (Kitcher 1984). Part of the resistance is due to historical inertia - philosophers have often characterized species as exemplars of natural kinds, and the individuality thesis presents a serious challenge to this useful characterization. Either species are not natural kinds, or philosophers need to radically revise their theories of natural kinds.
3.2 What kinds of individuals are species?
Let us accept the individuality thesis as a working perspective, as it is useful in setting up the next core issue facing systematists.[100] If the species category is the set of all individual species, what are the conditions for membership in this category? What kinds of individuals are species? Answering this question helps answer related questions: what are the relevant parts of a species, and when does speciation occur? The sheer number of answers provided to these questions is overwhelming, and constitutes what is typically referred to as the species problem, or debates over species concepts.
The species problem concerns what kinds of groups of organisms ought to count as being species. Even this characterization, though, presumes too much. For it might be that species are not groups of organisms, per se, but groups of populations, or parts of time-extended lineages (of populations, organisms, or some other genealogical group). Primary concerns for a species concept may range from discovery of some unit of evolution, to epistemological matters of specification (or both). Given space constraints, it will be most useful to take a broad look at two competing species concepts: the biological species concept (BSC) and the phylogenetic species concept (PSC).
The BSC (Mayr 1942, Coyne and Orr 2004) holds that species are interbreeding groups of populations, and that for an organism to belong to a species simply means it belongs to one of those interbreeding groups. Notice that there is some discrepancy as to whether or not the BSC implies actual or potential interbreeding is sufficient. Speciation occurs when a population becomes reproductively isolated from other parts of the species. Species are treated as units of evolution, bound together by a causal process that is likely to produce a unique group moving forward. The BSC is widely known and used, particularly by population biologists - in large part because looking at interbreeding groups coheres well with population biology models of evolution.
Systematists, on the other hand, more often prefer PSCs, of which there are several varieties. Unlike the BSC, PSCs are primarily concerned with groups of organisms with an exclusive shared history. Species, on a PSC account, are composed of organisms that are more closely related to each other than to any organism in a different species. Notably, interbreeding groups of organisms might not meet this criterion, leading to conflicts between PSC and BSC advocates, because the species boundaries given by one concept do not necessarily map on the boundaries given by another.
Constraining species to groups with unique and exclusive histories is useful for describing the pattern of evolution of taxa, and the distribution of characters across those taxa, both important inferences to be drawn from work in systematics (Baum 1992). When to mark a unique history is a matter of some debate among PSC advocates, ranging from very early (initial diagnosability) in a lineage split (Cracraft 1983) to very late (reciprocal monophyly) (Baum and Donoghue 1995) (see Wheeler and Meier 2000 for more on this and other debates).Many more species concepts have been proposed, and they have been widely discussed (e.g., Ereshefsky 1992a, Howard and Berlocher 1998, Wilson 1999b). Among the issues at stake is whether one ought to be a monist about species concepts, or instead accept a plurality of concepts as legitimate for biological research and theory (Sober 1984a, Ereshefsky 1992b) - or whether to simply reject the notion of species altogether (see especially Mishler 1999)! More recently, Kevin de Queiroz has argued that conflation of the problem of species concepts with the problem of species delimitation underlies much of the confusion around these debates (de Queiroz 2007; see also de Queiroz 1998 and Wilson 1999b). De Queiroz proposes that there is much more agreement surrounding species concepts than previously recognized, though much disagreement persists over criteria of speciation. It remains to be seen whether this position helps resolve disagreements, for either philosophers or biologists.
Notice that the species concepts described above reflect underlying theoretical commitments and the particular research interests of the biologists involved. This is not at all unique to species, but is a feature common to most concepts in science - particularly theoretical entities. More controversial is the philosophical significance of this fact. Kyle Stanford (1995), for instance, has asked whether we ought to adopt an anti-realist stance towards species.
3.3 Phylogenetic inference
The task of discovering and describing species turns on what kinds of individuals species are. What of the other major goal of systematics: determining and describing the phylogenetic relationships of species? Phylogeny is the pattern of common descent and is usually represented by phylogenetic trees
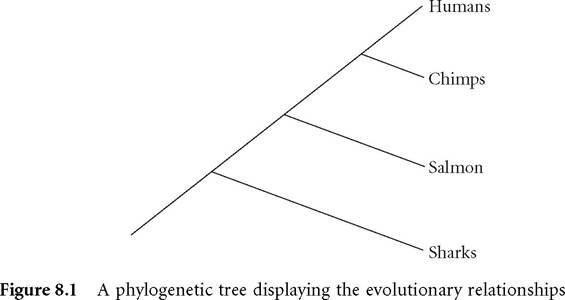
between taxa in terms of descent from shared ancestors. Tracing the lines from right to left traces the phylogenetic history of a taxonomic group, with more closely related groups sharing a more recent common ancestor. Thus, salmon are more closely related to chimps and humans than to sharks, by virtue of sharing a more recent common ancestor
(Figure 8.1). Following the branching lines in Figure 8.1 traces the pattern of evolution via common ancestry, and displays the phylogenetic relationships between these groups of species. Phylogenetic studies provide good grounds for evolutionary explanations and inferences - for example, we can appeal to Figure 8.1 to explain why humans and chimps share so many characters (they were passed down from a common ancestor), or to understand the claim that salmon are more closely related to chimps than to sharks (they share a more recent common ancestor).
Discovering phylogenies presents a problem familiar to philosophers of science - a special case of the problem of underdetermination of theory by evidence. For n number of taxa, there are exactly (2n - 3)!/2n-2(n - 2)! possible phylogenetic trees (Felsenstein 2004). In Figure 8.1, there are four taxa and, thus, 15 possible (rooted) trees, though only one that corresponds to the actual historical lineage. The number of possible trees increases exponentially with the number of taxa, such that for 10 taxa there are over 34 million possible trees! Worse, deep evolutionary history cannot be directly observed, and all of these possible trees are consistent with the data used to infer this history (where data are simply the distribution of characters across taxa). The challenge facing systematists is twofold: whether phylogenetic inference may be justified in light of such epistemic challenges, and which methods of phylogeny reconstruction allow such justification. As evidenced by the Wiens quotation above, contemporary systematists generally agree that inferring phylogeny is a legitimate (indeed, central) task in modern systematics, and philosophers and biologists have chronicled the emergence of this consensus (see especially Hull 1988).
More relevant to contemporary systematics are debates over which inferential methods provide a justified account of phylogeny. Initially, the phylogenetic technique of choice was parsimony analysis (Hull 1988, Sober 1988). Roughly, parsimony techniques select from among the possible trees the one proposing the fewest number of evolutionary events (Kitching, Forey, Humphries, and Williams 1998). Many leading cladistic theorists initially justified parsimony techniques by explicitly appealing to Karl Popper's falsificationism as a means of solving the problem of phylogenetic inference (Wiley 1975, Farris 1983). The most parsimonious phylogenetic tree was held to be a bold hypothesis of phylogeny, subject to falsification or corroboration depending on future analyses including new or additional data.
In the late 1970s, Joseph Felsenstein demonstrated that lineages of a certain shape were subject to a systematic error in parsimony analysis (Felsenstein 1978, 2004). One example of this is called long-branch attraction (named after the shape of the lineages; see Figure 8.2), in which taxa at the end of long branches are mistakenly grouped together by parsimony analysis, instead of with taxa with which they share a more recent common ancestor. This is not merely an operational problem, but a conceptual challenge to the falsificationist underpinnings of parsimony (Haber 2008). Felsenstein showed that as more data are included, the chance of long-branch attraction increases. This means that parsimony techniques are prone to rejecting hypotheses that correctly capture phylogenetic relationships while corroborating more parsimonious phylogenetic hypotheses that incorrectly group taxa - seemingly producing a systematic inferential error! Felsenstein proposed using statistical techniques (in particular Maximum Likelihood (or ML) methods) to avoid this problem.
Felsenstein's proposal was welcomed by some systematists (statistical phylogeneticists), but rejected by others, now typically called Cladists.[101] ML techniques were not seen as falling within a falsificationist framework, but instead were viewed as confirmationist. Cladists argued that the cost
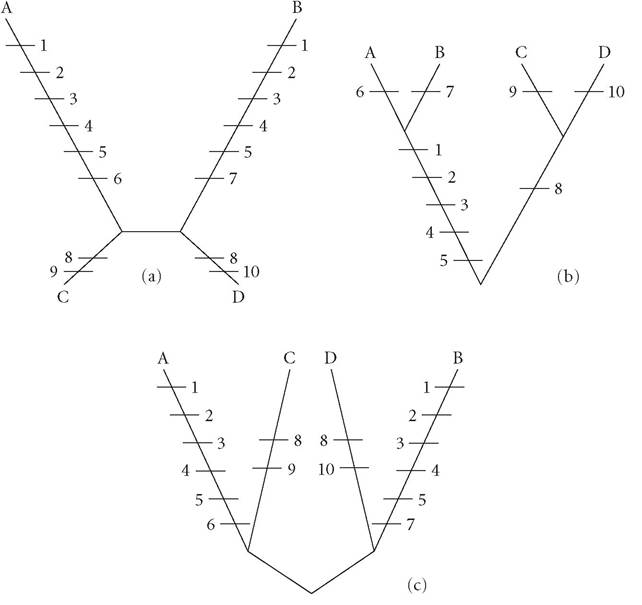
Figure 8.2 Long-branch attraction is a systematic error to which Maximum Parsimony Analysis is subject (see text). (a) is the actual historical pattern of evolution of taxa A, B, C, and D. The length of the branches corresponds to the amount of evolutionary change that has occurred along those branches, with marks representing evolutionary events. (b) is a parsimony analysis of the data from (a), whereas (c) is an ML analysis of that same data. Note that the phylogenetic relationships hypothesized in (b) are not isomorphic with the actual relationships in (a). The long-branch mistake occurs in (b) because parsimony analysis ranks possible phylogenetic hypotheses based on the number of proposed evolutionary events needed to account for all the characters exhibited by the taxa. For example, hypothesis (b) would receive a parsimony score of 10, whereas (c) would receive a parsimony score of 16
of adopting statistical techniques was to abandon good scientific protocol and a return of the problem of inference in systematics; only falsification- ism provided justified methods of surmounting the epistemic challenges facing systematists (e.g., Farris 1983). Thus the persisting debate between Cladists and statistical phylogeneticists is a philosophical disagreement, and not likely to be solved by data in particular cases. What counts as a success in such cases will itself be a matter of dispute. For example, it has been demonstrated that parsimony analysis can be fully construed using ML techniques (Tuffly and Steel 1997, Sober 2004), yet this remains unconvincing to cladists, who reject such characterizations of parsimony as illegitimate due to reliance on statistical approaches (Farris 1983).
4.
More on the topic Systematics:
- THE THEORY AND PRACTICE OF EMPIRE-BUILDING
- Unobtrusive Measures, Content Analysis, and Systematic Observation
- Any systematic review or synthesis of a vast amount of research studies within a certain spatial constraint is a daunting task.
- §104. Alchemy
- This chapter provides a systematic overview of the multidimensional measurement methodology of Alkire and Foster (2007, 2011a), with an emphasis on the first measure of that class: the Adjusted Headcount Ratio or M0.
- References
- Bayes's Theorem
- Conclusion
- The Concept of Identity: A Historical Perspective
- Multiple courses of corticosteroids