Anatomy of the Cell
Joanna M. Bassert
CHAPTER OUTLINE
INTRODUCTION, 46
OUR EARLY UNDERSTANDING
OF CELLS, 47
Evolution of Cells, 47
Size Limitations, 47
LOOKING INSIDE THE CELL, 48
Mammalian Cell Anatomy, 48
Cell Membrane, 52
Cytoplasm, 58
Nucleus, 67
LEARNING OBJECTIVES
When you have completed this chapter, you will be able to:
1.
Explain how scientists believe the earliest cells may have formed on earth.2. Differentiate between prokaryotic and eukaryotic cells.
3. List the early discoveries of the 17th, 18th, and
19th centuries that led to our modern understanding of cells.
4. List the physiologic factors that limit cell size.
5. Describe the molecular structure of the cell membrane.
6. Explain the importance of cilia and flagella.
7. List and describe the components of cytoplasm.
8. Describe the structure and functions of the cytoskeleton.
9. Describe the consistency and molecular components of cytosol.
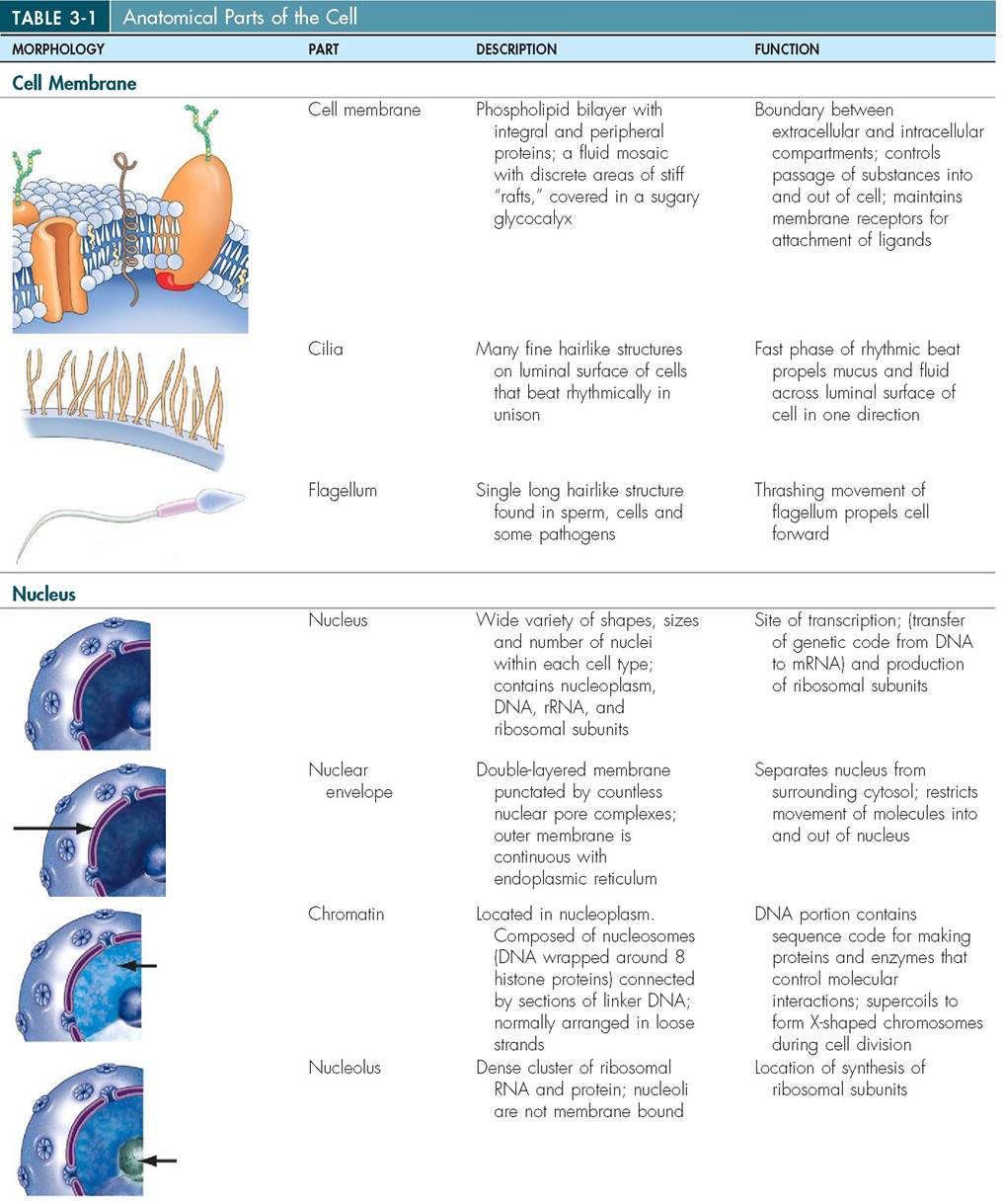
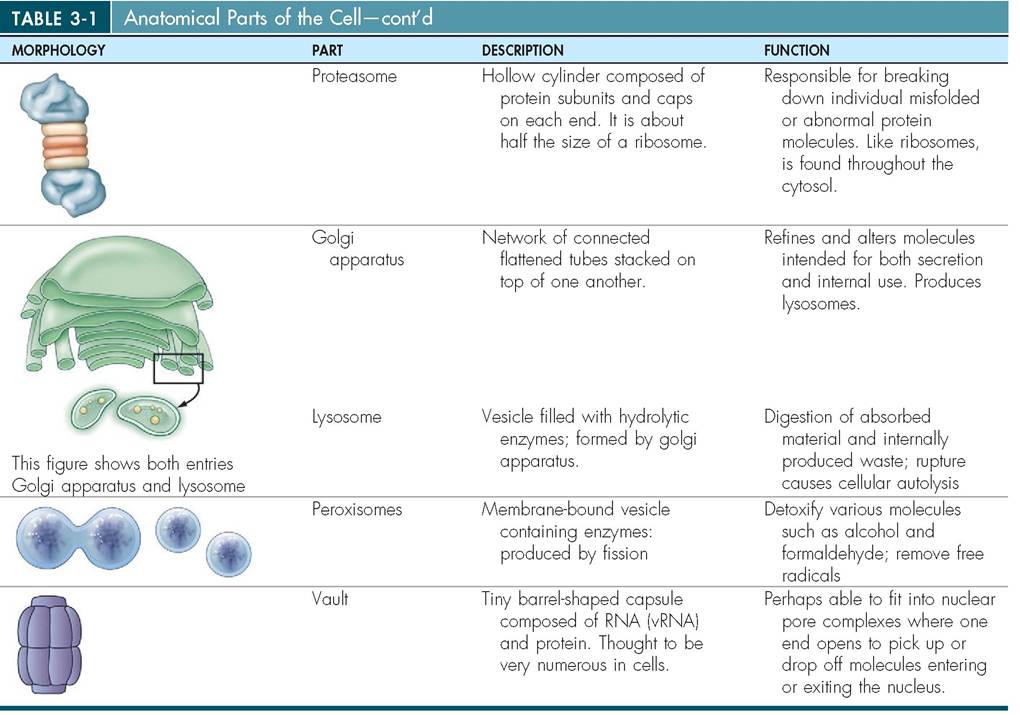
10. Describe the structure and functions of each of the cellular organelles, inclusions, and vesicles.
11. List the parts that make up the nucleus; for each part, explain its structure and function.
12. Compare and contrast the molecular structure of DNA and RNA.
13. Define the term “chromatin” and explain the relationship between chromatin and chromosomes.
VOCABULARY FUNDAMENTALS
Adenine ahd-eh-nιn
Autolysis aw-tohl-uh-sihs
Basal body bas-sahl boh-de
Catalyze kaht-ih-lιz
Caveolae ka-ve-eh-le
Cell sehl
Cell adhesion molecule sehl ahd-he-shuhn mohl-uhl-kyool
Cell-mediated immune response sehl me-de-a-tehd ih-myoon reh-spohns
Cell membrane sehl mehm-bran
Centriole sehn-tre-ol
Chemical signaling kehm-ih-kuhl sihg-nahl-ihng
Chromatin kro-mah-tihn
Chromosome kro-mo-som
Cilia sihl-e-ah
Cristae krih-ste
Cytoplasm sι-to-plahz-uhm
Cytosine sι-to-sen
Cytoskeleton sι-to-skehl-ih-tuhn
Cytosol sι-tuh-sahl
Deoxyribonucleic acid (DNA) de-ohck-se-rι-bo-noo-kla- ihck ah-sihd
Electron microscopy e-lehck-trohn mι-kraw-sko-pe Endoplasmic reticulum ehn-do-plahz-mihck reh-tihck-u-luhm
Enzyme ehn-zιm
Eukaryote yoo-kear-e-ot
Flagella flah-jehl-ah
Fluid mosaic floo-ihd mo-za-ihck
Genetic material jeh-neht-ihck mah-teer-e-ahl
Globular protein glohb-yoo-lar pro-ten
Glycolipid glι-ko-lihp-ihd
Glycoprotein glι-ko-pro-ten
Golgi apparatus gol-je ahp-uh-raht-uhs
Guanine gwah-nen
Histone hihs-ton
Immunization ihm-u-nι-za-shuhn
Inclusion ihn-kloo-shuhn
Integral protein ihn-teh-grahl pro-ten
Intermediary fiber ihn-tar-me-de-ear-e fi-bar
Keratin fiber kear-ah-tihn fi-bar
Ligand lι-gahnd
Light microscopy lit mι-kraw-skδ-pe
Lipid bilayer lihp-ihd bi-la-ar
Lysosome li-so-som
Mathias Scheiden mah-thi-uhs shi-dehn Matrix ma-trihks
Membrane receptor mehm-bran reh-sehpt-ar Microfilament mι-krδ-fihl-ah-mehnt Microtrabeculae mι-krδ -truh-behck-u-la Microtubule mι-krδ-too-bul
Mitochondria mι-tδ-kohn-dre-ah
Multicellular organism muhl-ti-sehl-u-lar ohr-gah-nihz-uhm
Myofibril mι-δ-fi-brihl
Neurofilament nar-δ-fihl-ah-mehnt
Nuclear envelope noo-kle-ar ehn-veh-lδp Nucleoli noo-kle-δ-lι
Nucleoplasm noo-kle-δ-plahzm
Nucleosome noo-kle-δ-sδm
Nucleus noo-kle-uhs
Organelle ohr-gah-nehl
Organism ohr-gah-nihz-uhm
Osteocyte ohs-te-δ-sit
Peripheral protein puh-rihf-ar-ahl pro-ten
Peroxisome par-ohcks-ih-sδm
Plasma membrane plahz-mah mehm-bran
Plasmalemma plahz-mah-lehm-ah
Prion pri-ohn
Prokaryote prδ-keor-e-δt
Proteasome pro-te-ah-sδm
Protoplasm pro-tδ-plahz-uhm
Raft rahft
RNA: Ribonucleic acid rι-bδ-noo-kla-ihck
ah-sihd
Robert Hooke rohb-art hook
Scanning electron microscopy skah-nihng eh-lehck-trohn mι-kraw-skδ-pe
Scrapie skra-pe
Specialization spehsh-uh-lah-za-shuhn
Spindle apparatus spihn-duhl ahp-uh-raht-uhs
Spindle fiber spihn-duhl fi-bar
Theodor Schwann the-uh-dohr shwahn
Thymine thi-mihn
Tonofilament toh-nδ-fihl-ah-mehnt
Transmission electron microscopy trahnz-mihsh-uhn eh-lehck-trohn mι-kraw-skδ-pe
Uracil yar-ah-sihl
Vault vahlt
Virus vi-ruhs
INTRODUCTION
The cell is the remarkable basic unit of living things.
It can exist alone as a single, free-living plant or animal, or it can combine with other cells to form elaborate complex organisms, such as trees, horses, and people. The cell is dynamic and performs all of the functions by which life is defined. It has a metabolism, can grow, develop, reproduce, adapt, respire, become influenced by outside stimuli, maintain a stable internal environment, and convert food into usable energy. Each cell carries vital genetic material that governs its own development, metabolism, and specialization. A great diversity is seen in the appearance of cells. They can be small, biconcave red blood cells; long, thin myofibrils; or octopus-like osteocytes. Each form reflects its own specialized function (Figure 3-1). In a multicellular organism, such as a dog, cells have differentiated and become grouped into specialized tissues that work collaboratively to sustain life for the animal as a whole. These tissues, as well as the systems they form, are the focus of anatomy and physiology, but it is important to remember that their functional unit is the cell. It is in the cell that molecular messages are transmitted and received, electrical impulses generated, oxygen absorbed, and energy manufactured. Thus we must first learn about cells before we can understand the anatomy and physiology of the tissues and systems they compose.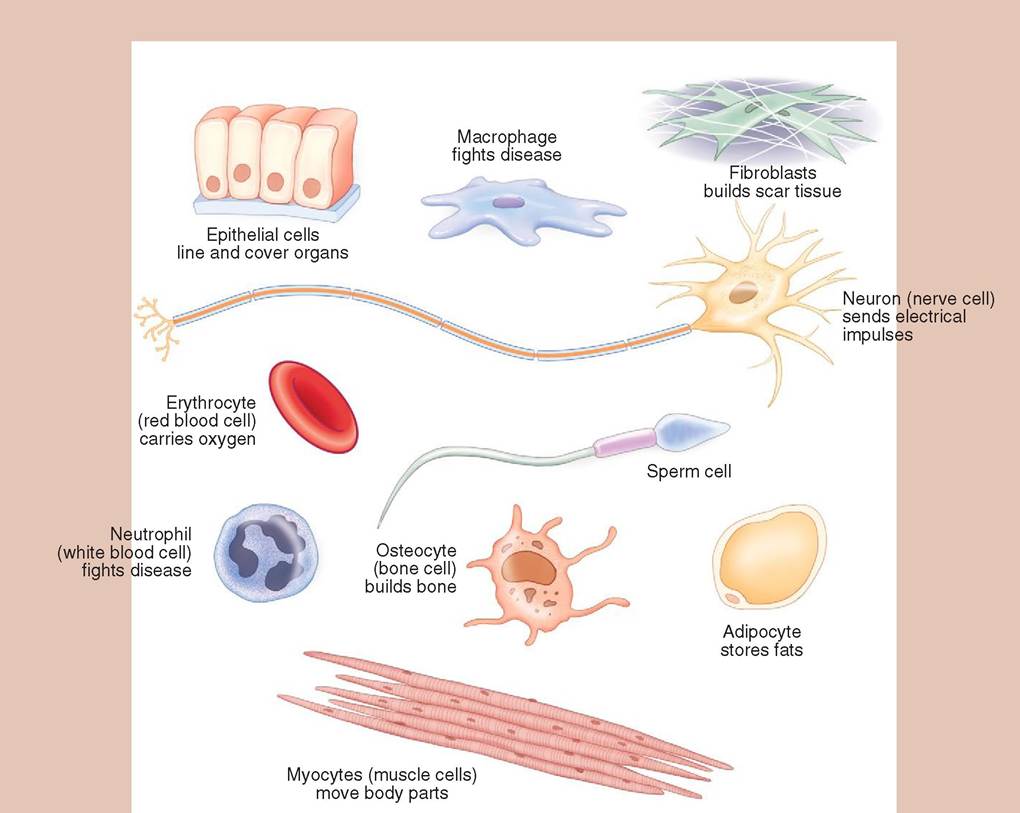
FIGURE 3-1 Exampl es of cell types. Differentiation and specialization of cells in the multicellular organism has led to a diverse array of cell types. The shape and size of the cell are related to its function.
OUR EARLY UNDERSTANDING OF CELLS
EVOLUTION OF CELLS
The first cells are thought to have evolved in the massive oceans of our primitive earth about 3 billion years ago. Jolted by the fierce electrical energy from frequent lightning storms and by the intense, unabated radiation from the sun, the three molecules that made up the primitive atmosphere— methane gas (CH4), water (H2O), and ammonia (NH3)— were forced to collide and split apart.
The first organic molecules, similar to amino acids, were thought to have formed in this tempestuous environment. Clustering into heavy droplets, these molecules are thought to have been washed Irrnri the atmosphere by driving rains into the warm, shallow seas below. There, proteins, lipids, and carbohydrates ehvicohlved, w in turn arranged themselves over time into sophisticated, organized structures to become the first cells.Tfihese primitive cells resembled present-day bacteria and contained a single strand of deoxyribonucleic acid (DNA), which floated freely in gelatinous protoplasm. The cells did not possess a nucleus and were therefore called prokaryotes, which means before nucleus. For 1.5 billion years prokaryotes were the only forms of life on this planet. Eukaryotes, meaning true nucleus, developed later and are found in all multicellular organisms today. Eukaryotic cells are charac- tyerized b a distinct nucleus, in which the DNA has com- ibtihned w protein to form chromosomes. These, in turn, are surrounded by a protective nuclear envelope, wh,clι, life a igouna,rd stat monitors the flow of molecules in and out of tuhcelenus.
SIZE LIMITATIONS
The size of most animal cells is restricted to a range of 10 to 30 μm in diameter because of the relationship between the surface area and the volume of a cell. Small cells have smaller nutritional requirements than large cells but have a proportionately large surface through which they can absorb the substances they need. Thus a small cell with a proportionately large surface area will be able to complete its metabolic functions more rapidly and efficiently than a large cell with a relatively small surface area. In other words, if cells were the size of basketballs, they would not be able to take in nutrients fast enough to feed themselves and would die.
A second factor limiting cell size is related to the governing capability of the nucleus. A single nucleus can control the metabolic activity of a small cell better than a large one.
Also, the more active a cell is, the greater are its metabolic needs. Therefore it is not surprising that some very large cells or cells that are very active, such as cardiac and skeletal muscle cells, have two or more nuclei. Also, these cells are long and thin, thereby creating a larger surface area through which nutrients can be absorbed. The combination of increased nuclei and expanded surface area enables muscle cells to function at a very high metabolic rate./ Testyourself 3-1
1. What are the basic cellular functions that define life?
2. Describe the series of events that scientists believe led to the formation of the first cells on earth.
3. What is the difference between a prokaryote and a eukaryote?
4. Why are cells not the size of watermelons?
LOOKING INSIDE THE CELL
MAMMALIAN CELL ANATOMY
Today with the aid of sophisticated microscopes, scientists have been able to look inside cells and focus on their minute, fantastic internal world (Box 3-1). For over 3 billion years, cells have evolved into diverse shapes and have taken on a wide range of specialized functions within multicellular organisms. Despite these changes and the morphological diversity that evolved among them, there are three essential structures found in all mammalian cells
BOX 3-1 An Introduction to Microscopy
In the 1600s, a Dutch scientist named Anton van Leeuwenhoek found microscopic creatures in pond water. He was able to make this exciting discovery using a single-lens microscope that he had fabricated himself. The Englishman Robert Hooke also made his own microscope, which allowed him to observe the tiny units that make up cork. These early, homemade tools were the first light microscopes and marked the beginning of three and a half centuries of impressive exploration in the frontiers of microspace.
Light Microscopy
Light microscopy works by using visible light to see tiny structures. Tissues and other items being examined are sliced or spread out into thin layers, affixed to glass slides, and stained.
When placed on the stage of the microscope, as shown in the drawings below, the tissues can be examined through the ocular lens. Light comes from underneath the slide and is concentrated on the tissue by the condenser. The effect of this backlighting is dramatic and colorful. Pictures taken of microscopic objects using a light microscope are called light micrographs and are found throughout this and other anatomy texts. Use of stains, including fluorescent stains, has markedly improved the visibility of microscopic structures using light microscopy.Electron Microscopy
As its name implies, electron microscopy uses electrons rather than light to create an image. An electron beam generated by an electron gun is directed with precision toward the specimen by magnets. Some of the electrons pass through the specimen, but some of them bounce off and are deflected to the sides. Electron beams are not visible to humans, and the generation of images in electron microscopes is therefore dependent upon an electron detector, which translates the pattern of incoming electrons into a readable image. The electron microscope is so powerful that it can make extraordinarily minute intracellular structures visible, including the organic molecules that compose cells. Interestingly, there is no color at this level of magnification, because the structures being examined are as small as, or smaller than, the wavelength of the colors we humans can see. However, computer enhanced coloration is used to augment the black and white world, creating images with greater clarity and definition.
There are two types of electron microscope: transmission electron microscopes (TEM), which generate two-dimensional images; and scanning electron microscopes (SEM), which generate three-dimensional images. Transmission electron microscopy works by sending a beam of electrons through a specimen in the same way that light microscopy sends a beam of light through a specimen. Thus the specimen used in TEM, like light microscopy, must be very thin so that the electron beam can pass through it.
Once the electrons pass through the specimen, they strike phosphorescent particles on a viewing screen, which release photons (visible light). In this way, the screen literally glows with the microscopic image.Specimens in scanning electron microscopy are first "sputtered" or covered with tiny particles of gold before they are examined. Inside the microscope, an electron beam is generated and is scanned back and forth over the specimen, hence the term scanning electron microscope. The electrons from the beam are deflected at the surface and secondary electrons are displaced from the surface as well. These primary and secondary electrons are detected by a sensor that relays the
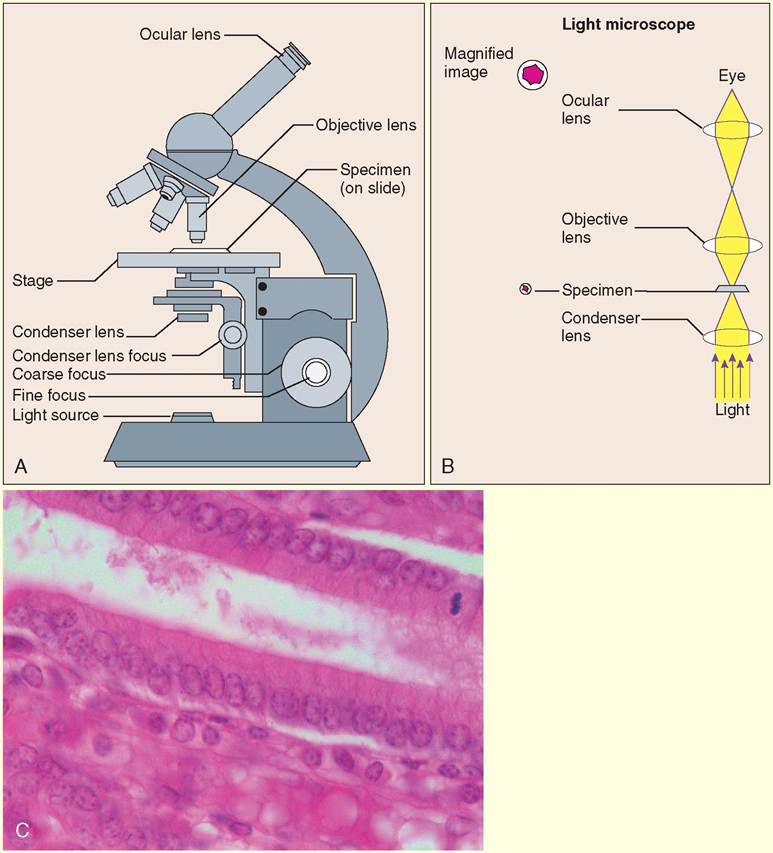
Light microscopy (LM). A, The light microscope uses a light source to illuminate very thin tissue samples, which are put on glass slides and placed on the microscope's stage. The objective lens and ocular lens together create the magnification needed to enlarge the image. B, Specimens under examination are backlit by visible light, which is focused on the stage by the condenser. C, The simple columnar epithelial cells that cover the folded lining of the abomasum (stomach) in a steer come to life with LM. Stain is absorbed by the nuclei, making them appear purple. Cytoplasm and the cell membranes are various shades of pink. (A, B, From Thibodeau GA, Patton KT: Anatomy & physiology, ed 6, St Louis, 2007, Mosby; C, courtesy Joanna Bassert.)
signal to an amplifier. The magnified image is subsequently displayed on a monitor.
Atomic Force Microscopy
An atomic force microscope (AFM) generates a threedimensional image of incredibly small structures using a minute sharp-tipped probe attached to a delicate cantilever (arm). The tiny probe meticulously scans the surface of specimens, allowing the forces between the tip and the sample surface to deflect the cantilever. This deflection is recorded by a computer, which generates a digital image. Some of the forces that can cause deflection include capillary forces, chemical bonding,
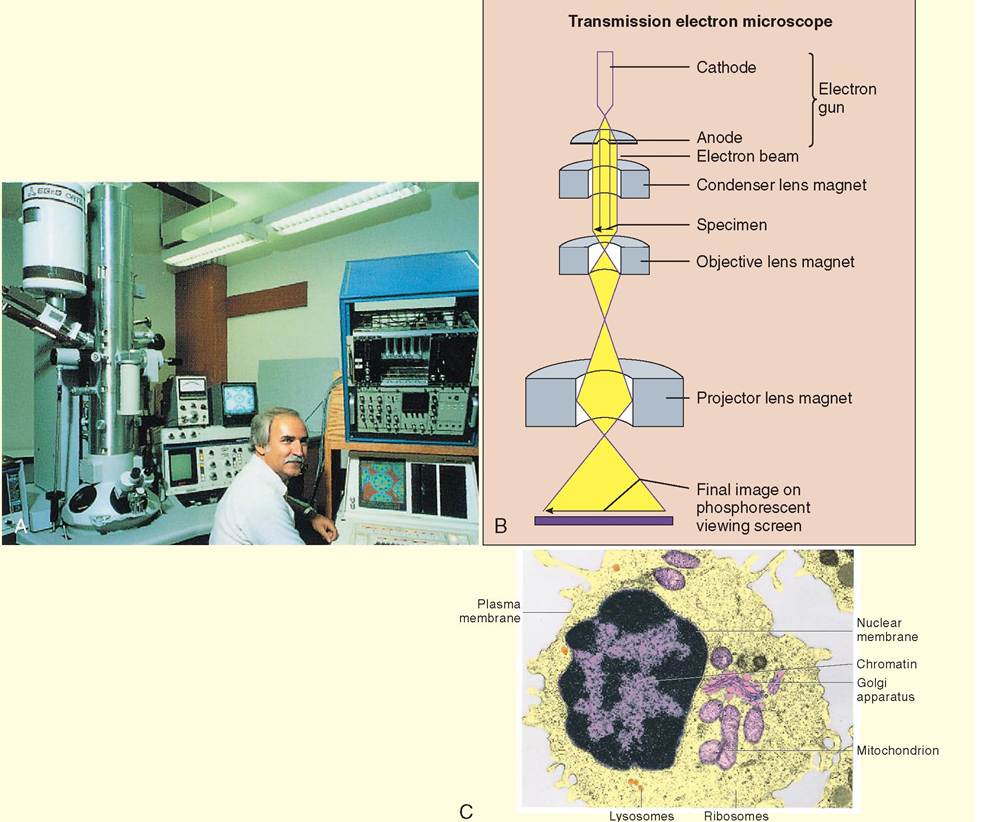
Transmission electron microscopy (TEM). A, An electron microscope and the associated monitors fill an entire room. During examination of specimens, it is helpful if the room is dark. B, An electron beam is generated by the electron gun. The direction of the beam is focused on a specimen by condenser magnets. Electrons that pass through the specimen bombard phosphorescent particles on the viewing screen, where the image is seen. C, A transmission electron micrograph of a mammalian cell. Far greater levels of magnification are possible with electron microscopy than light microscopy. (From Thibodeau GA, Patton KT: Anatomy & physiology, ed 6, St Louis, 2007, Mosby.)
electrostatic and magnetic forces, as well as mechanical contact forces. In most AFMs, a feedback mechanism is employed by the computer to adjust the tip-to-sample distance so that a constant force is maintained between the tip and the sample. In this way, the tip and sample are prevented from banging into one other causing damage to either the sample or the tip. The tip of the probe is often covered with an array of special coatings depending upon what forces are being investigated. For example, a gold coating is used to trace covalent bonding in organic molecules, diamond coatings are used to increase the wear resistance of the probe and magnetic coatings are used to evaluate magnetism on the surface of a
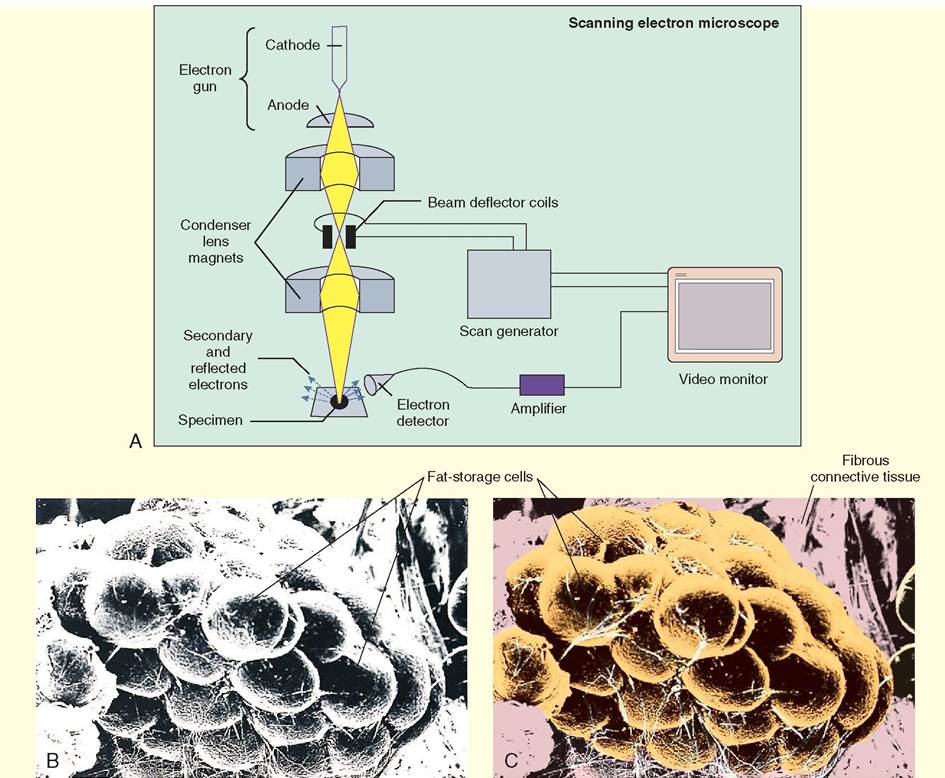
Scanning electron microscopy (SEM). A, An electron beam is scanned back and forth over a specimen. The electrons that bounce off the specimen, and secondary electrons that come from the surface of the specimen, are detected, amplified, and transcribed into an image, which is visible on a video monitor. B and C, SEM brings adipose tissue to life in threedimensional images. Electron micrographs can be enhanced with color by computers to add a greater level of detail and clarity. Notice that the pink added to the background helps to delineate the fibrous connective tissue from the fat cells. (From Thibodeau GA, Patton KT: Anatomy & physiology, ed 6, St Louis, 2007, Mosby.)
specimen. The probe can scan the surface of specimens in a static (or contact mode), or via a dynamic (noncontact) mode in which the probe is vibrated. The results are magnificent three-dimensional images on a scale far smaller than previously thought possible. The use of AFM technology has enabled scientists to look more deeply inside cells and to visualize the minute anatomical structures that are necessary for life. Thus, AFM has led to a greater understanding of the cause of some diseases that have devastated both humans and animals.
Continued
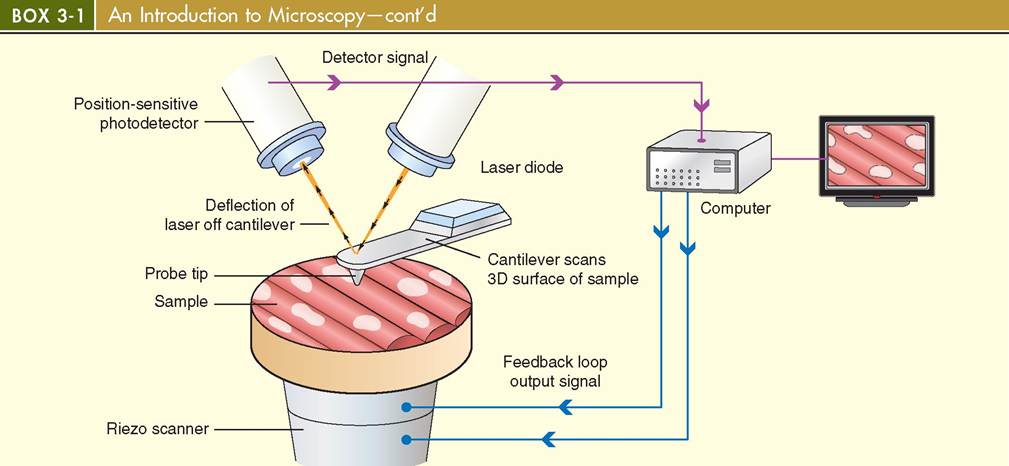
Atomic Force Microscopy (AFM)
Atomic force microscopy (AFM) uses a minute probe and cantilever system to scan the surfaces of specimens. A laser and a position-sensitive photodetector translate the minute movements of the probe into data that are subsequently used to generate a three-dimensional image on a computer screen.
regardless of their shape, size and function. These structures are:
• The cell membrane (plasmalemma)
• The cytoplasm:
• Cytosol
• Cytoskeleton
• Organelles
• The nucleus
All cells are separated from their environment by a cell membrane, which is also known as a plasma membrane or plasmalemma (plaz-ma-LEM-a; lema meaning husk). Everything inside the cell membrane, other than the nucleus and genetic material, is known as the cytoplasm (cyto means “cell”). Cytoplasm includes: cytosol, a colloidal, jamlike protoplasm that is highly structured and composed of proteins, electrolytes, and metabolites; a flexible cytoskeleton; and complex structures called organelles. Organelles, like the organs in our own bodies, work collaboratively to carry out necessary metabolic functions (Table 3-1). As already mentioned, all mammalian cells are eukaryotic and therefore possess a nucleus, which contains vital genetic material in the form of chromosomes. Figure 3-2 is an illustration of a composite cell that includes many of the important structures students are asked to learn and study. Keep in mind that, in reality, no single cell includes all of these features and cells have a diverse array of shapes and sizes depending upon their function within the body. Also, be aware that nuclei have many shapes also and that many of them are not positioned in the center of the cell.
CELL MEMBRANE
The cell membrane acts as a flexible, elastic barrier between the inner cytoplasm and the outside environment (Figure 3-3). It includes many infoldings and outpouchings that provide extra surface area, and it is continually removing and recycling different segments of itself, updating surface receptors and renewing its sticky outer coating. Like our own skin, the cell membrane is capable of self-repair, but if it is damaged to the extent that intracellular contents are released, the cell quickly dies. The cell membrane governs the movement of atoms and molecules in and out of the cell. Although the actual consistency and complexity of the cell membrane are based largely on the function of the cell, cell membranes usually consist primarily of protein (55%) and phospholipids (25%) but also include quantities of cholesterol (13%), miscellaneous lipids (4%), and carbohydrates (3%).
MEMBRANE STRUCTURE
Because of its surprising thinness (about 60 to 100 angstroms), the cell membrane is not visible using light microscopy. Under the transmission electron microscope, however, the cell membrane appears as two thin, dark layers with a seemingly empty space between them. At the molecular level, the cell membrane is composed of two layers of phospholipid molecules arranged so that the hydrophilic heads are on the outside and the hydrophobic, fatty acid tails are on the inside. This is called a lipid bilayer. Proteins that are suspended in this bilayer can move easily throughout the membrane to create a constantly changing mosaic pattern

known as the fluid mosaic. Most lipid-soluble materials, such as oxygen and carbon dioxide molecules, pass through the membrane with ease, whereas ionized and water-soluble umcohlecules, s as amino acids, sugars, and proteins, which are not lipid soluble, do not readily pass through.
The membrane also contains cholesterol molecules that wedge themselves between phospholipids and help to stabilize the membrane. In this way cholesterol not only prevents the lipids from aggregating, which helps to keep the membrane fluid, but it also adds to the oily nature of the internal layer, which increases the membrane's impermeability to water-soluble molecules.
eTlhl e c membrane is composed of a wide variety of
important structural and globular proteins, which are responsible for the membrane's special functions. Compact rgolotebiunlsar p may occur either on the cell surface or inside the lipid bilayer. Those that occur within the bilayer laerde cal integral proteins.hTese molecules span the entire
fwidth o the membrane and may create channels through which other molecules can pass. Some integral proteins form selective passageways that permit only particular substances trnoteer o exit the cell, whereas others create pores,ucsh channels within the protein molecule that allow substances such as water to pass through with no resistance. Pores are svcearttered o the surface of the cell and make up approximately 0.2% of the cell's surface area.
Other globular proteins form peripheral proteins, which are bound to the inside or outside surfaces of the cell mem- bhreane. T internal peripheral proteins are more restricted in their movement than integral proteins, because they are cohftedn atta to portions of the internal cytoskeleton or to
txhpeoseed parts of some integral proteins. Peripheral proteins sometimes act as enzymes to catalyze specific chemicaI reactions and may be involved in the mechanics of changing the cell’s shape, an event that is quite dramatically seen, for euxrainmgple, d the contraction of a muscle cell.
rThe inne and outer layers of the cell membrane are dif- foemrent fr one another. Proteins that reside on the inner fsurface o the membrane, for example, may be bound to components of the cytoskeleton, keratin fibers, or periph- eoal proteins, whereas proteins and lipids on the outer layer are attached to sugar groups. These glycoprotein (sugar and protein) and glycolipid (s^ar and phospholipid) molecules are the principal components of the “sugar coating” that covers the surface of cells. This coating is called the glycocalyx. Life the pattern of stripes on a zebra or the fingerprints
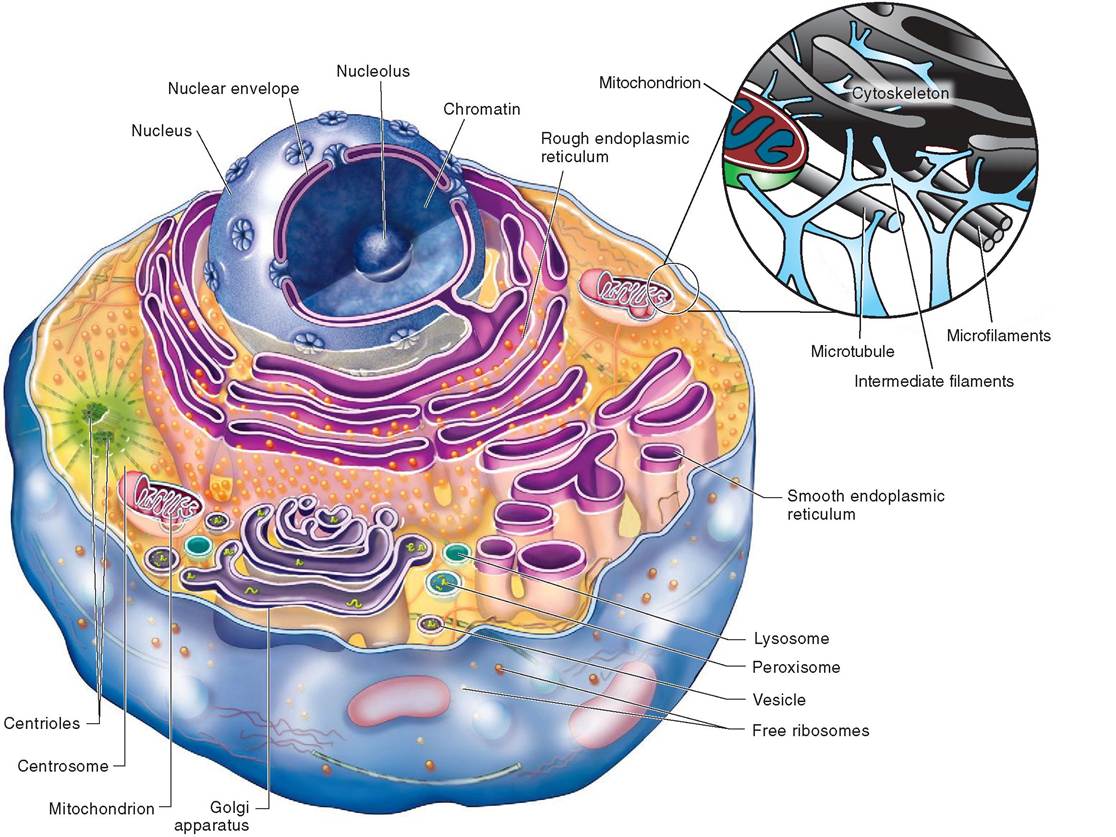
FIGURE 3-2 Composite example of a mammalian cell. (Modified from Patton KT, Thibodeau GA: Anatomy & physiology, ed 8, S Louis, 201 3, Mosby.)
Oiici human hand, the glycocalyx is unique. It provides improved cel 1-to-cel 1 adhesion and represents an important biological marker for intercellular recognition and for the interactions between the cell and antibodies or the cell and viruses. ⅞e interaction between the glycocalyx and extracellular molecules may bring about changes in the membrane aonssdibply in the activity of the cell as a whole.
The glycoca 1yx is composed of two families of molecules: cell adhesion molecules and membrane receptors. Cell adhesion molecules (CAMs) are sto^lky! glycoproteins that cover the surfaces of almost all of the cells in mammals; they allow the cells to bond to extracellular molecules and to each other. Tliese molecules are also important in helping cells move pnaest o another and in signaling circulating cells, such as white blood cells, to areas of inflammation or infection.
Membrane receptors are integral proteins and glycoproteins that act as binding sites on the cell surface. Some of them play a vital role in cell-to-cell recognition, a process called contact signaling. THs is particularly important during the cell-mediated immune response and assists bacteria and viruses in finding preferred target cells. Membrane receptors are also involved in a process called chemical signaling. (Hormones, neurots^^^rntters, and other chemical messengers called ligandsieonbcdific t sp binding sites on cell surfaces Once bound to the cell membrane, ligands can bring about a change in the cell's activity. Some ligands act as enzymes trcotivaate o inactivate a particular cellular activity.
Molecules that make up the fluid mosaic are not evenly distributed across the cell membrane. Some areas, for example, are stiffer and more rigid than others. As molecules irnteehact w o another within the fluid mosaic, rigid areas eonf sdely packed phospholipids, cholesterol, and protein form plaquelike structures called rafts wit bin the cell mem- bhreayne. T appear to move on the surface of the cell much
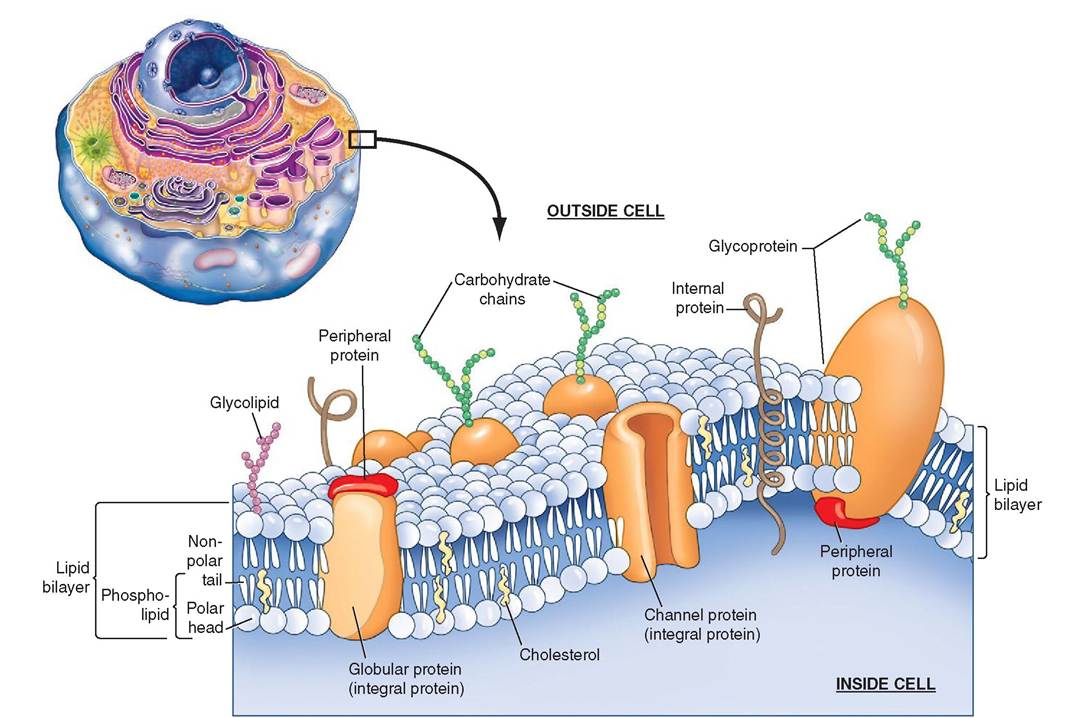
FIGURE 3-3 Cell membrane. The cell membrane is composed of a lipid bilayer. Proteins help govern the movement of atoms and molecules in and out of the cell. (Top left, Modified from Patton KT, Thibodeau GA: Anatomy & physiology, ed 8, St Louis, 201 3, Mosby.)
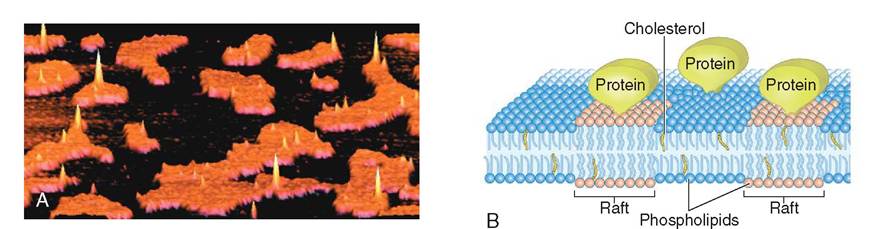
FIGURE 3-4 A, An image generated by an atomic force microscope shows rigid orange-colored rafts surrounded by darkly colored phospholipids on the surface of a cell. Proteins are visible reaching upwards into the spaces above the rafts. B, An illustration of the molecular structure of a fluid mosaic membrane containing rigid rafts. Notice that the rafts are rich with cholesterol molecules and protein. These molecules cling to one another, forming a single unit that moves through the phospholipid bilayer, much like a raft on water. Rafts are important in membrane-bound organelles as well as in the cell membrane, where they facilitate molecular interactions and functions. (A, Used with permission: Henderson R et al: Lipid rafts: feeling is believing, Physiology 19(2):39-43, © 2004 Int Union Physiol Sci/Am Physiol Soc.)
like a log floating on water. Rafts are important in helping to organize functional areas on the cell surface, in initiating cytokinesis during cell division, and in forming depressions on the cell surface that may lead to involution and vesicle formation. With use of atomic force microscopy, rafts are visible as discrete plaques in a sea of phospholipids on the cell membrane (Figure 3-4).
CAVEOLAE. Caveolae are small (50 nm) invaginations of the plasma membrane. Often the caveolae pinch off and
CLINICAL APPLICATION
Insulin is an important ligand that binds to specific receptors on the surface of cells, particularly liver, fat, and cardiac and skeletal muscle cells. Insulin stimulates glucose uptake by triggering the movement of glucose transporter protein 4 (GLUT4) from an intracellular storage compartment to the plasma membrane. Once inside the cell, glucose is used to manufacture adenosine triphosphate (ATP), a vital energy source that fuels metabolic functions. Without adequate amounts of ATP the cell may not be able to function properly, can weaken and may even die. In many patients with type 2 diabetes (also known as diabetes mellitus), the number of insulin receptors on the cell surface is decreased. This is a physiologic response to excessive fat stores in the animal and is one form of insulin resistance. With fewer insulin docking sites in the cell membrane, diminished amounts of glucose are brought inside the cell. Paradoxically, the cell begins to starve even though the level of glucose outside the cell is very high (too high).
migrate inside the cell to form tiny vesicles. Depending upon the type of cell, caveolae can form single vesicles or clusters of them, like grapes on a vine or tiny rosettes. The word caveolae means “little caves” and pertains to their appearance as minute invaginations on the cell surface. Caveolae are produced only from rafts in the plasma membrane that contain the integral protein caveolin. Cells that lack caveolin also lack caveolae. Three types of caveolin have been identified thus far (caveolin-1, caveolin-2, and caveolin-3). All of them possess a similar loop structure that looks as though it was inserted into the cell membrane like a hairpin. Caveolin-1 and caveolin-2 have been found in the cell membranes of endothelial, fibrous, and adipose cells, while caveolin-3 has been found in striated and smooth muscle cells.
Many cellular functions have been associated with caveolae including endocytosis, transcytosis, and contact signaling. Caveolae with specialized receptors can capture extracellular material and bring it into the cell. In endothelial cells, for example, caveolae are known to shuttle substances such as low density lipoprotein (LDL—the infamous “bad” cholesterol) from the luminal side of the cell to the basal side (Figure 3-5). As the caveolae discharge their LDL cargo, the space behind the endothelium fills with cholesterol, forming a plaque that over time will thicken and narrow the lumen of the vessel. These atherosclerotic plaques can obstruct blood flow and lead to heart disease and stroke.
FLAGELLA AND CILIA
Flagella and cilia are extensions of the plasma membrane that extend into the extracellular space. They are energetic, motile “hairs” that are structurally identical but function differently from one another. Cilia and flagella are composed of nine pairs of microtubules that encircle a central pair of microtubules. Both cilia and flagella originate from a pair of centrioles, called basal bodies, which are located at the periphery of the cell, just under the plasma membrane.
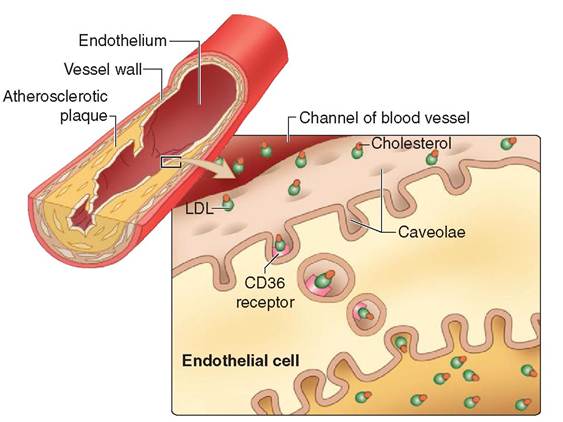
FIGURE 3-5 Caveolae. The endothelial lining of blood vessels is rich with caveolae. The caveolae fitted with CD36 receptors will attract low density lipoproteins (LDL), also known as the 'bad' cholesterol. The caveolae involute and pinch off from the plasma membrane, forming transport vesicles that carry the cholesterol to the other side of the endothelial cell. Here the LDL is discharged into the space behind the endothelium. Over time, a plaque forms in the wall of the vessel, narrowing the lumen and increasing the likelihood of a blockage. If the vessel blocks completely, the tissues that it normally supplies with nutrients and oxygen will die. Tissue death due to hypoxia is called an infarct. If an infarct occurs in the heart, it is called a myocardial infarct, which is a common cause of a heart attack. (From Patton KT, Thibodeau GA: Anatomy & physiology, ed 8, St Louis, 2013, Mosby.)
During their formation, cilia and flagella grow outward from the basal bodies and exert pressure on the plasma membrane.
Cilia occur in large numbers on the exposed surface of some cells (Figure 3-6). They are shorter than flagella and measure only about 10 μm long. They move synchronously, one after the other, creating waves of motion that propel fluid, mucus, and debris across the cellular surface. Cilia are best known for their important functions: (1) in the upper respiratory tract, where they propel bacteria and mucus away from the lungs; and (2) in the oviduct, where their beating motion pulls the expelled egg away from the ovary and into the opening of the oviduct.
Flagella generally occur singly and are significantly longer than cilia. They are typically attached to individual cells and propel the cell forward by undulating. Flagella move cells through fluid, whereas cilia move fluid across cell surfaces. The tail of a sperm cell is an example of a flagellum. A sperm is the only mammalian cell that is propelled by a single flagellum, although many disease-causing organisms are propelled in this manner.
Cytoplasm
The cytoplasm is the inner substance of the cell, excluding the nucleus. Initially, seen under light microscopy, it appears as a nondescript bag of gel with a few opaque speckles. Now, with the increased use of electron microscopy, cytologists
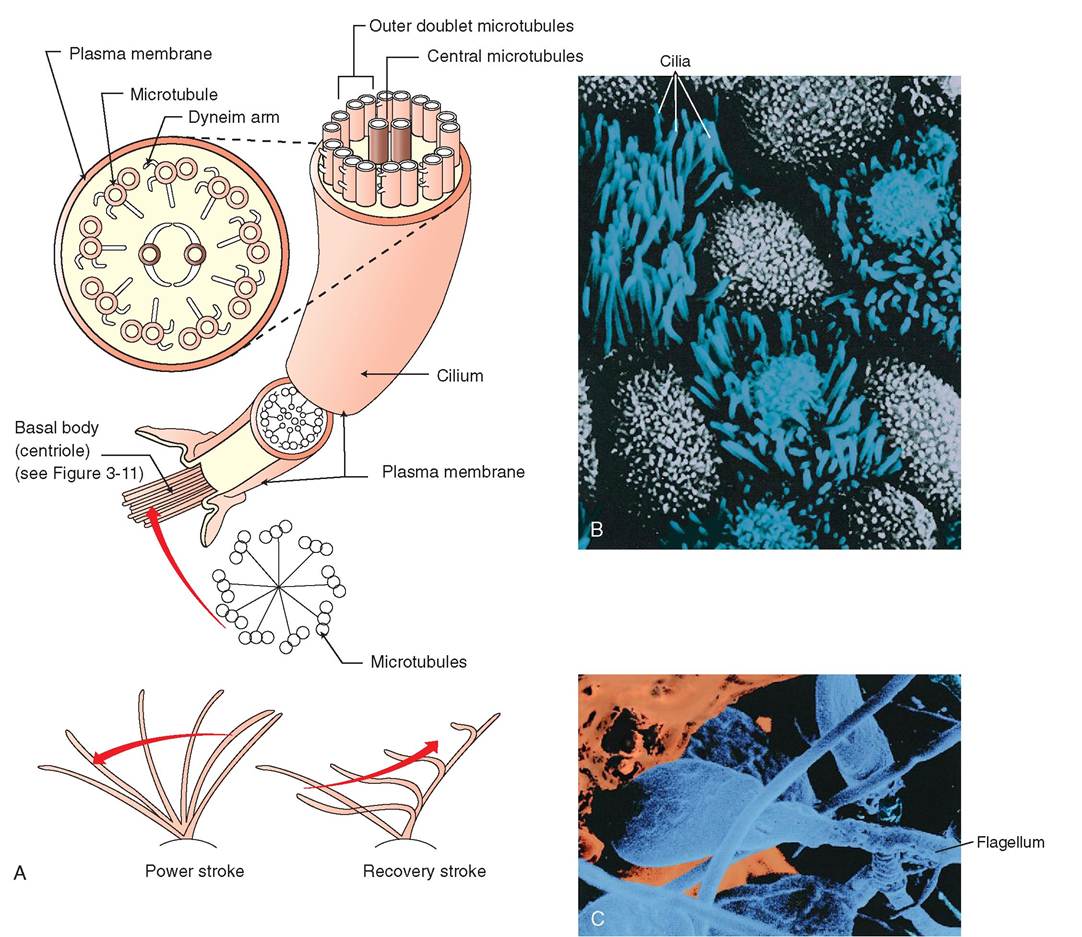
FIGURE 3-6 Cilia and flagella. A, Cilia and flagella are composed of nine pairs of microtubules that surround a central pair. Cilia have a fast, powerful, and synchronized stroke in one direction. In the trachea, this helps to propel particles away from the lungs toward the mouth where they can be coughed up and swallowed. Unlike cilia, which are numerous on the surface of some c∈jlls, flagella occur singly. B, A scagning electron micrograph of cilia on the apical surface of epithelial cells in the lung airways. C, A sr^nraig electron micrograph of sperm. The single elongated flagellum enables the sperm cell to move rapidly through the female repeoducti.e tract. (B, Coertesy Charles Flickinger, University of Virginia. C, From Thitadeau GA, Patton KT: Anatomy & & physiology, ed 8, St Louis, 201 3, Mosby.)
TEST YOURSELF 3-2
1. Name three Structuresthatallmammaliancells possess.
2. Draw a pistrre or tHιa∣erbd bSayer.WhicS partishydraphobicsndwhicS partis hydrophilic?
3. WSat typei ot eroihinare fount inthmcallmembrane?
4. WSere are tleseproto inslocaSad and what aceineir funoiionsh Addth eorto mour drawing.
5. WSat are tha mamcuiarcemponeetsof rafts ardwAartoladoraftselayia tha life of a cell?
6. WSat isnhdg∣anocWos asdwhar importaet ro Id Hoen itplay ic CeHuIarinteraction?
7. WCSat areCAMs CinA whata? they do?
8. WhSat aremametaae reaoptorsoed iigacds, aad whaSrold totheyploy it tie HeaIte ot tie coll?Hawdoes obesity affect cell memhrane renedtars tar inshlinO
9. WSat are sevectas ance whatra∣edo theye∣ay isthhcsllmembrane?
ho. Htrw ore ciθa and HegeHo different?
11. WSinS are taunO mare namman∣o in mammalian cells: cilia ar flagella?
have the ability to visualize minute internal structures that define the complex internal workings of the cell. The principal components of cytoplasm are the cytosol, cytoskeleton, organelles, and inclusions.
CYTOSOL
The fluid of the cell is called cytosol. It is a viscous, semitransparent liquid composed of dissolved electrolytes, amino acids, and simple sugars. Proteins are also suspended in the cytosol and give it its thick, jellylike consistency. These proteins are mostly enzymes that are important in the metabolic activities of the cell.
Cytoskeleton
Like the skeleton in our bodies, the cytoskeleton is a threedimensional frame for the cell; but unlike our bones, it is neither rigid nor permanent (Figure 3-7, A and C). It is a flexible, fibrous structure that changes in accordance with the activities of the cell. It gives support and shape to the cell, enables it to move, provides direction for metabolic activity, and anchors the organelles. Three different types of fiber comprise the cytoskeleton, all of which are made of protein. They are microtubules, intermediate fibers, and microfilaments (Fir^re 3-7, B). These fibers are not enclosed in a membrane.
The thickest fibers are the microtubules, which are long, hollow tubes that grow out from the cell center near the nucleus. They form secure “cables” to which mitochondria, lysosomes, and secretory granules attach. Proteins that act as motors move the attached organelles along the microtubule from one location in the cell to another. Because microtubules act as the railroad tracks for organelle travel, they can be easily disassembled and then reassembled to form new paths or to take on new direction. They appear, for example, in greater numbers during cell division to assist in the separation of chromosomes and organelles. Microtubules are composed of a pair of spherical molecules called tubulins, which are linked together into a spiral chain. The spiral shape provides strength and flexibility to cilia and flagella, as well as to the cell as a whole.
Intermediate fibers are woven, ropelike fibers that possess high tensile strength and are able to resist forces pulling on the cell by acting as internal guy wires. These fibers are the toughest and most permanent element of the cytoskeleton. They are composed of different proteins, depending on the function of the cell, and often take on different names depending on the type of cell in which they are found. In epithelial cells, for example, intermediate fibers are composed primarily of keratin and are known as tonofilaments or keratin filaments, whereas in nerve cells they are known as neurofilaments.
Microfilaments are located near the cell surface on the cytoplasmic side of the plasma membrane and are arranged in bundles and meshworks. They are composed of the contractile protein actin and, together with the motor protein myosin, play a key role in the cell's ability to change shape, break apart during cell division, and form outpouchings and involutions. In most cells, microfilaments are assembled where and when they are needed. Their position and quantity within the cell vary depending on the cell's activity. In muscle cells, however, the microfilaments are permanent, highly developed myofibrils, which shorten to cause muscle contraction.
Some cytologists believe that minute microtrabeculae exist as a fourth component of the cytoskeleton. These fibers are thought to form a lattice that interconnects larger cyto- skeletal elements, suspends free ribosomes, and gives cytosol its jamlike consistency.
Centrosome. A important region of the cytoskeleton, located near the nuclear envelope, is called the centrosome. The centrosome is responsible for the coordination of building and breaking down microtubules in the cell and is composed of several parts: centrioles, pericentriolar material, and asters (Figure 3-8). The centrosome helps manufacture microtubules, the thickest and strongest fibers in the cell. Microtubules are not only important parts of the cytoskeleton but also key elements in cell division and the formation and function of cellular extensions such as cilia and flagella. In these ways, the centrosome is vital to cellular life.
A pair of small cylinders, called centrioles, is found in the central portion of the centrosome. Each centriole is composed of nine triplets of microtubules arranged like a pinwheel around a hollow core. Long rods of microtubules, called asters, radiate away from the centrioles, forming anchors to the larger centrosome. In the center of this region, a concentrated amorphous collection of proteins forms the pericentriolar material (PCM), where formation of microtubules is initiated.
In preparation for cell division, centrioles must first duplicate themselves; each one forms a “daughter” centriole that is oriented perpendicular to the “mother” centriole. Mother and daughter centrioles are linked to one another by a thin interconnecting fiber. The mother centriole is equipped with an array of molecular appendages that help anchor the newly formed microtubules that emerge from the PCM. As the fibers lengthen, the two centriole teams move slowly apart from one another, holding fast to the fibrous ends of the elaborate construction that ultimately becomes the spindle apparatus.
The centrosome plays an important role in the formation of microvilli, cilia, and flagella. Refer to the section in this chapter on the cell membrane for more information about these microtubular cell extensions.
ORGANELLES
Organelles, or “little organs,” are membrane-bound structures within the cytoplasm that possess specialized cellular functions. The membranes of the organelles are similar in composition to those in the plasma membrane but do not have glycocalyx coatings. In this way, each organelle is separated from the surrounding cytosol and is able to maintain its own internal environment. This compartmentalization is crucial for effective metabolic processes, because it enables the cell to separate and control various molecular
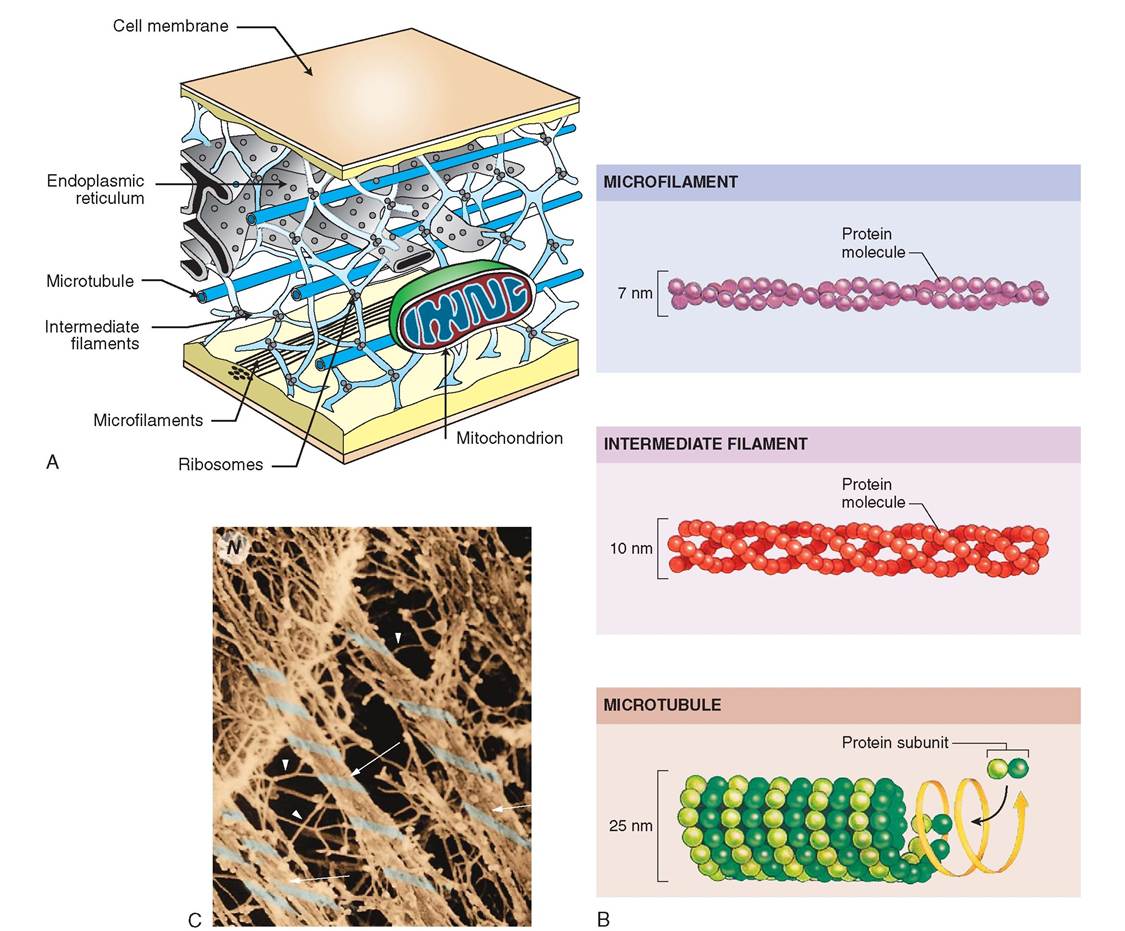
FIGURE 3-7 Cytoskelet on. A, The cytoskeleton is an important structural frame for the cell. It is strong but flexible and can be dismantled in some regions and rearranged as needed. B, Left: Cytoskeleton is composed of three different types of fiber: microtubules, intermediate fibers, and microfilaments. Middle: Electron micrographs of each type of fiber. Right: Light microscopy reveals parts of the cytoskeleton using fluorescent stains to illuminate specific molecules. C, Color-enhanced scanning electron micrograph showing cytoskeleton. Note that the microtubules (arrows) are thicker than the minute intermediate filaments (arrowheads). (B, Top, Courtesy I. Herman, Tufts University; middle, Courtesy E. Smith and E. Fuchs, University of Chicago; bottom, courtesy G. Borisy, University of Wisconsin, Madison.)
interactions, which are the basis for food absorption, energy production, and excretion.
MITOCHONDRIA. Among the largest of the organelles is the mitochondrion (plural, mitochondria). This is known as the powerhouse of the cell because it produces 95% of the energy that fuels the cell. In the mitochondria, large nutrient molecules, such as glucose, are processed and broken down into smaller ones, which can be used intracellularly to fuel most metabolic processes (Figure 3-9). It is also where respiration takes place: oxygen is consumed, and carbon dioxide is excreted. Numerous biochemical reactions occur in the mitochondria, such as amino acid and fatty acid catabolism, respiratory electron transport, oxidative phosphorylation, and the oxidative reactions of the citric acid cycle. (See Chapter 17 for more detailed descriptions of these metabolic processes.)
Active cells, which have high energy demands, have greater numbers of mitochondria within their cytoplasm than inactive cells. Heart muscle cells, for example, have far
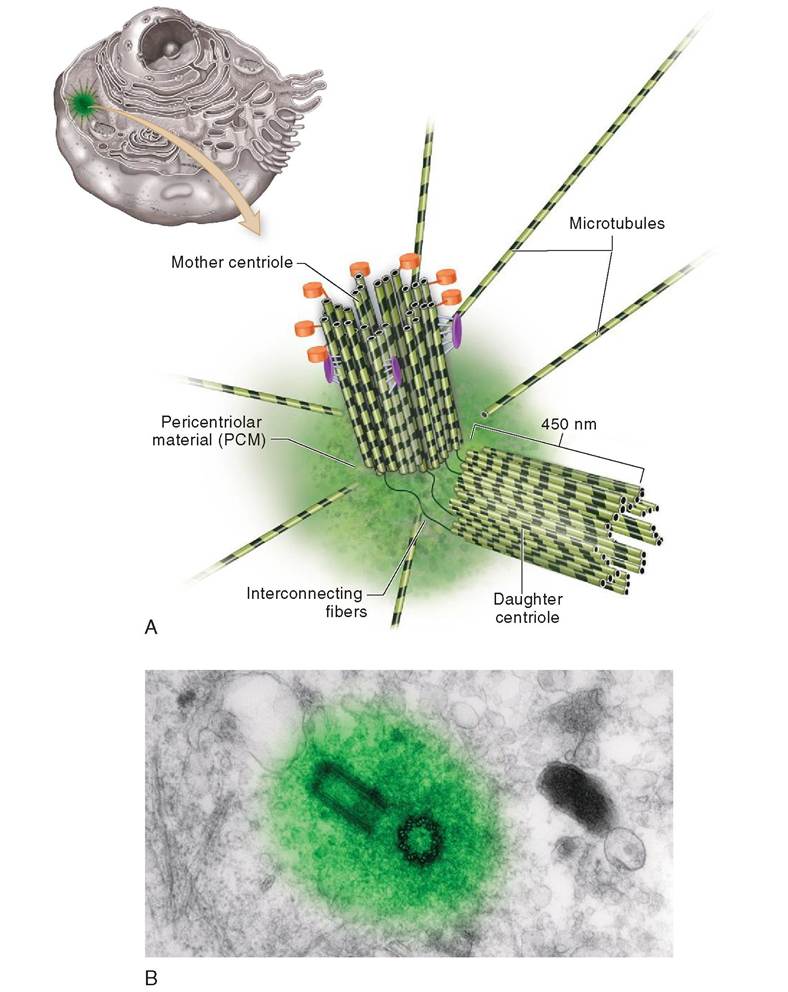
FIGURE 3-8 Centrosome. A, A mother and daughter pair of centrioles form the focus of the centrosome. Each centriole is composed of nine triplets of microtubules arranged around a central axis, much like a pinwheel. Minute fibers connect the centrioles to one another while microtubules called asters radiate outward from the pericentriolar material. B, A transmission electron micrograph (TEM) of a centrosome. (B, Photograph by Dr. Conly L. Rieder, East Greenbush, NY.)
more mitochondria than relatively inactive endothelial cells. When cellular requirements for energy increase, the mitochondrion divides by pinching itself in half via a process called fission. Both halves subsequently grow to normal size. In addi^n, mitochondria tend to congregate in areas of the cell where greater amounts of energy are required, such as at the base of a flagellum.
Mitochondria contain the DNA, RNA, and enzymes necessary to make protein, but they provide themselves with only 13 of the proteins required for their metabolic functions; the nucleus provides the remaining 50. Thus most of trhoeteipn needed by the mitochondria is produced elsewhere in the cell and is later taken up by the mitochondria.
Mitochondria may take on a variety of shapes but tend to lbliepteical or round. They can move throughout the cell laonndgactaen e or change shape with ease. Mitochondria are enclosed by two membranes: the outer one is smooth and featureless, and the inner one involutes dramatically, forming
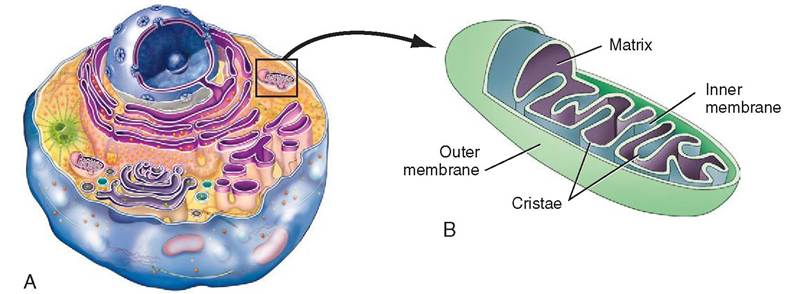
FIGURE 3-9 Mitochondrion. A, The mitochondrion is the powerhouse of the cell. Using oxygen, it produces 95% of the energy that fuels the cell. B, Transmission electron micrograph of a mitochondrion. Note the convolutions of the inner membrane, which create increased workspace for the manufacturing of adenosine triphosphate (ATP). (Modified from Thibodeau GA, Patton KT: Anatomy & physiology, ed 6, St Louis, 2007, Mosby.)
shelflike folds called cristae. These folds increase the internal working area and allow greater contact between the cristae and the enzyme-rich liquid, called the matrix, which fills the spaces between the cristae. In addition to containing vital enzymes, the matrix is composed of calcium ions and the substrates required for metabolic reactions. Additional enzymes are available in the form of small particles that are found attached to some of the cristae. Because the cristae are the site of ATP production, it is not surprising that active mitochondria possess more cristae than inactive ones.
The DNA and RNA found in mitochondria are similar to those found in bacteria but are quite different from those found in the nucleus and cytoplasm. Mitochondria are thought to have originated as independent, bacteria-like organisms billions of years ago that later moved into the bodies of unicellular plants and animals, developing a symbiotic relationship with them.
RIBOSOMES. The most common and smallest organelle in the cell is the dark-staining ribosome (Figure 3-10). It is composed of two globular subunits, which fit together like cupped hands. These subunits contain protein and a specific type of RNA, known as ribosomal RNA (rRNA). Although only 25 μm in diameter, the ribosome is an important site for protein synthesis. Soluble protein intended for intracellular use is manufactured on ribosomes that are evenly distributed freely throughout the cytoskeleton. Protein intended for use in the plasma membrane or meant for cellular export, on the other hand, is synthesized on ribosomes attached to the endoplasmic reticulum. Ribosomes are flexible in their abilities to attach and detach from membranes and to move freely within the cell. Thus they can move back and forth between the cytoskeleton and endoplasmic reticulum, depending on the type of protein they are making. When manufacturing protein, the ribosomes assemble the amino acids into long chains using specific instructions that are determined by the cell's genetic material. In this way, a wide range of proteins, such as cellular enzymes, hormones,
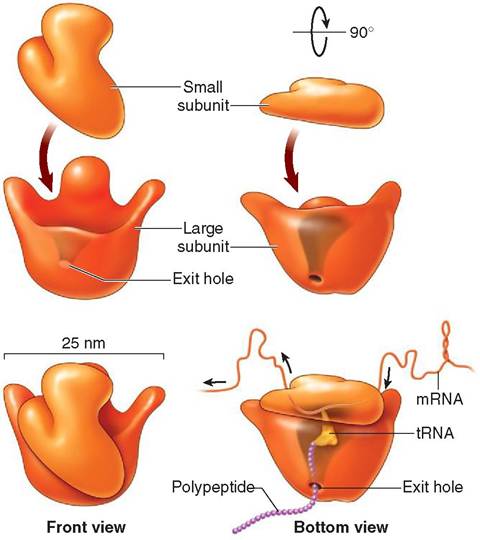
FIGURE 3-10 Ribosome. Each ribosome is composed of one large and one small subunit. When joined together, these two subunits form one ribosome. The small subunit attaches to a strand of messenger RNA (mRNA). Amino acids are carried to the ribosome by transfer RNA (tRNA). The amino acids are assembled in order according to the recipe in the genetic code. As a chain of amino acids forms and lengthens, it leaves the ribosome via an exit hole in the bottom of the large subunit. When the polypeptide is fully formed, it will be released from the ribosome. (From Patton KT, Thibodeau GA: Anatomy & physiology, ed 8, St Louis, 201 3, Mosby.)
collagen, and mucus, may be created, based on the needs of the cell and of the organism as a whole.
ENDOPLASMIC RETICULUM. The endoplasmic reticulum (ER) is a series of flattened tubes stacked on one another and bent into a crescent shape. Its surface area is enormous and may be 30 times larger than that of the plasma membrane. The walls of the ER are composed of a single lipid bilayer and are continuous with the membranes of the nucleus.
The two types of ER are rough, which has ribosomes on its surface, and smooth, which lacks ribosomes. Rough ER is involved in the production of protein, which is assembled by the ribosomes. These newly manufactured protein molecules are moved inside the ER into passageways known as cisternae (sis-TUR-ne; a reservoir of water). Here the proteins are modified before being moved on to the Golgi apparatus for further modification and packaging. Smooth ER, which is connected to rough ER, is active in the synthesis and storage of lipids, particularly phospholipids and steroids, and is therefore seen in large quantities in gland cells. In liver cells, it may also function to eliminate drugs and break down glycogen into glucose. The proportion of smooth to rough ER varies depending on the synthetic activities of the cell.
GOLGI APPARATUS. The Golgi apparatus, like the ER, is composed of stacks of flattened, crescent-shaped cisternae (Figure 3-11). Small, spherical transfer sacs from the ER containing newly manufactured proteins are received by the Golgi apparatus. These sacs fuse with the membrane at the ends of the Golgi apparatus and dislodge their contents into the Golgi cisternae. The protein molecules are then moved from stack to stack through the Golgi body, where they are modified. For example, sugar groups, which the Golgi apparatus manufactures, may be added to the proteins to form glycoproteins. When the modified proteins reach the outermost layer of the Golgi's flattened tubes, they are packaged into small spherical vesicles. The vesicles are formed as they pull away from the cisternae and venture out into the cytosol, where they travel to other parts of the cell, particularly the cell membrane. Thus the Golgi body acts as a modification, packaging, and distribution center for molecules destined either for secretion or for intracellular use. It also functions in polysaccharide synthesis and in the coupling of polysaccharides to proteins to create glycoproteins found on the cell surface.
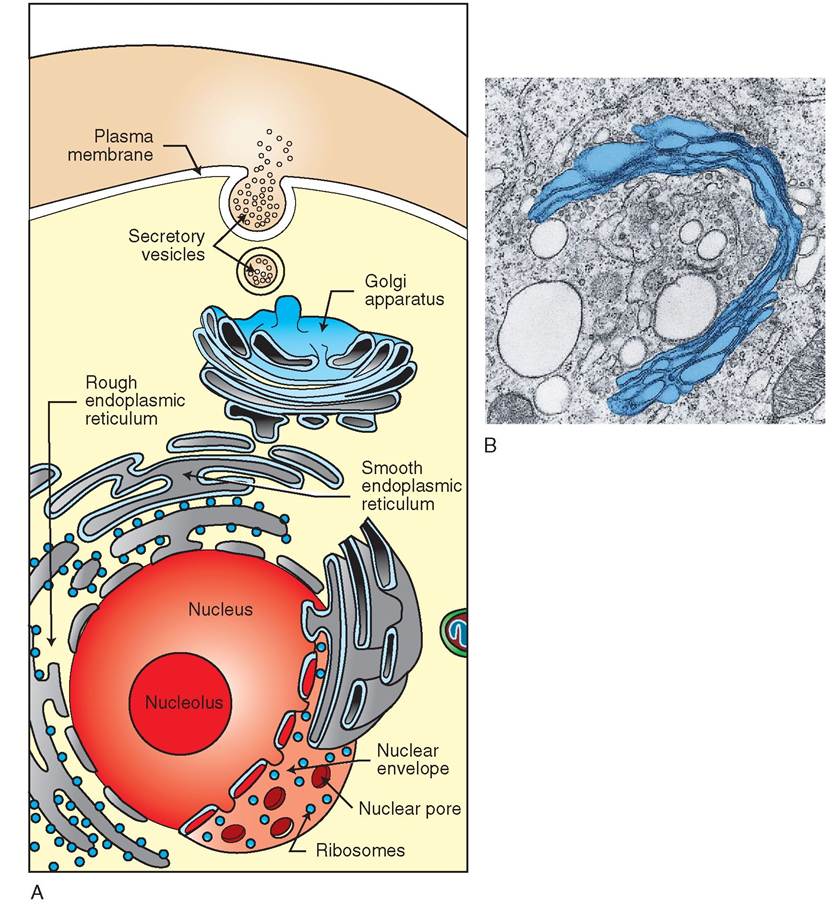
FIGURE 3-1 1 Golgi apparatus and endoplasmic reticulum. A, Endoplasmic reticulum (ER) is continuous with the nuclear envelope and is divided into two types: rough ER, which is covered by ribosomes, and smooth ER, which does not have ribosomes. Molecules produced by ER are transported to the Golgi apparatus, where they are modified and packaged into vesicles. Vesicles transport molecules to other regions of the cell. B, A transmission electron micrograph of the Golgi apparatus. (A, Courtesy Charles Flickinger, University of Virginia. B, From Thibodeau GA, Patton KT: Anatomy & physiology, ed 8, St Louis, 2013, Mosby.)
LYSOSOMES. The lysosome is a specialized vesicle formed by the Golgi apparatus (Figure 3-12). It contains powerful enzymes enclosed in a single, protective membrane, which fuses with other vesicles carrying nutrients, microbes or aged cellular parts. Both the ER and the Golgi apparatus are skilled in policing the cytosol for unused and damaged protein, organelles, and cellular debris. Their membranes reach out into the cytosol and envelope this unwanted material. Subsequently, the vesicles fuse with lysosomes, which dump their digestive enzymes into the vesicles to break down the contents. A small residual body is all that is left behind, carrying amino acids and other molecular subunits that can be recycled to form new molecules or neatly expelled from the cell if not needed. It is not surprising, therefore, that the lysosome is considered the “stomach” or “garbage disposal” vessel of the cell. Its
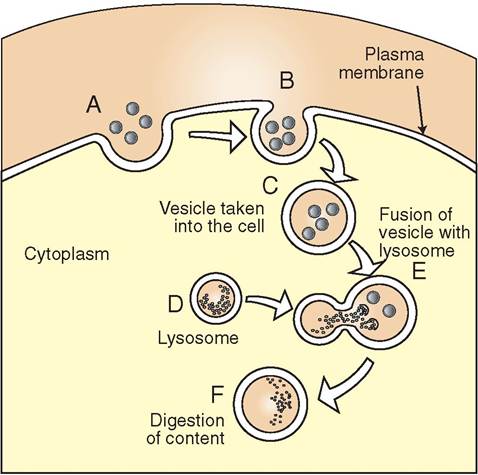
FIGURE 3-12 Lysosomal action. A, Material from outside the cell is drawn into a forming vesicle. B, The plasma membrane surrounds the material and pinches off to form a vesicle. C, The vesicle transports material to internal regions of the cell. D, A lysosome approaches the vesicle. E, The lysosome fuses with the vesicle and dumps digestive enzymes into it; these break down the material. F, The contents of vesicle are digested and transported to other regions of the cell.
principal responsibilities are to break down nutrient molecules, aged organelles, and cellular debris and to destroy phagocytosed microinvaders. Lysosomes also remove accumulations of protein that might otherwise obstruct normal cellular processes.
When cells die, the lysosomes within them are triggered to burst open and release their caustic enzymes into the cytosol; these enzymes immediately begin to dismantle and digest the various organelles and nuclear components of the cell. The process of self-digestion is called autolysis. The organism as a whole recycles the used parts of the dead cell to create new cells or to help maintain existing ones. Not surprisingly, as cells age, the number of lysosomes within them increases.
Lysosomes may also release their enzymes outside the cell to assist with the breakdown of extracellular material. During the process of bone remodeling, for example, osteocytes use lysosomes to help break down and remove unnecessary bone. This process is seen radiographically (in an x-ray film) as bone resorption, with a decrease in opacity of the affected area. In addition, lysosomal digestion is responsible for decreasing the size of body tissues, for example after parturition, with shrinkage of the uterus, and in the atrophy of muscles in paralyzed animals.
PROTEASOMES. The proteasome is a small cylindrical structure composed of multiple protein subunits. Like the lysosome, it assists with the breakdown and removal of misfolded and unwanted protein in the cell. Unlike the lysosome, however, which can digest large amounts of cellular debris, the proteasome can only break down one protein at a time.
The proteasome is barrel-shaped and composed of a stack of flattened protein subunits with a hollow core that extends down its center (Figure 3-13). Regulatory molecules form
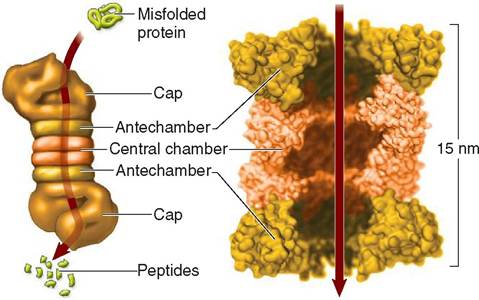
FIGURE 3-13 Proteasome. Vital to the health of cells, the proteasome disposes of damaged, old or unwanted protein in cytoplasm that otherwise might build up and obstruct normal metabolic activity. The tiny organelle is a little more than half the size of a ribosome and is made of protein subunits with caps at each end. Following the route indicated by the arrows, a misfolded protein enters the hollow core of the proteasome where it is broken down into short peptide chains. The peptide chains exit the proteasome at the opposite end, and are further broken down into amino acids, which are used to make new proteins.
caplike structures at each end of the proteasome. These molecules control the entry and release of the proteins that it processes.
Before a protein can enter the proteasome, a short- chained protein called ubiquitin is attached to the misfolded protein. The ubiquitin then “pulls” the protein into the central core of the proteasome. As the protein passes through the cap, the protein is unfolded to prepare it for processing in the central chamber where it is broken into short peptide chains 4 to 25 amino acids in length. These short peptides are subsequently expelled from the distal cap into the cytosol where they are further broken down into amino acids that are recycled to make new proteins.
Scientists have learned that normal functioning of protea- somes is essential for healthy cells. In diseases such as Parkinson's disease in humans, for example, accumulations of misfolded proteins kill cells in the brain that regulate muscle function, because the proteasome system is not functioning properly.
PEROXISOMES. Like lysosomes, peroxisomes are membranous sacs containing enzymes that are found throughout the cell. But unlike lysosomes, which are formed by the Golgi apparatus, peroxisomes are formed by vesicles pinching in half via fission or by pinching off from the ER. They commonly occur in liver and kidney cells and are important in the detoxification of various molecules. Peroxisomes contain enzymes that use oxygen to detoxify a number of harmful substances, including alcohol and formaldehyde. They also assist in the removal of free radicals, which are normal products of cellular metabolism but can be harmful to the cell in large quantities because they interfere with the structure of proteins, lipids, and nucleic acids. Peroxisomes carry two major types of enzyme: peroxidases, which assist in the conversion of free radicals to hydrogen peroxide, and catalases, which reduce hydrogen peroxide to water.
VAULTS. A new tiny organelle has recently been discovered, called a vault (Figure 3-14). Its minute barrel-shaped structure with tapered ends is reminiscent of a straight “crescent” dinner roll, but unlike dinner rolls, vaults are hollow inside. Vaults are thought to be extremely numerous throughout the cell and may play a role in transporting molecules to and from the nucleus. They appear to be attached to microtubules in the cytoskeleton and are thought to act as transportation pods, sliding rapidly from one end of the cell to another. Their tiny size enables them to dock in nuclear pore complexes where they can open into a rosette shape that echoes that of the nuclear pore complex. In this way, they can open one end to pick up or drop off ribosomal subunits or large molecules. They are made of protein and a small amount of RNA. This RNA is called vault RNA (vRNA or vtRNA for short).
TEST YOURSELF 3-3
1. What are the principal components of cytoplasm?
2. What is cytosol and what kind of molecules are found in it?
3. What is the centrosome and what important roles does it play in the life of the cell?
4. What is the cytoskeleton and what is its function?
5. How many types of fiber make up the cytoskeleton? Can you name them? How do they function differently?
6. Draw a picture of each of the eight organelles described earlier.
7. How does each of these organelles function within the cell?
INCLUSIONS
Inclusions are packaged units of metabolic products or substances that the cell has engulfed. They may be delineated by a surrounding, single-layer membrane, as seen in secretory granules, vacuoles, and vesicles, or they may be non-membrane bound inclusions, such as lipid droplets and fat globules. Vacuoles are larger than vesicles but are otherwise identical in structure. They are often filled with water and solutes that are transported to and from the cell surface. Some vesicles act simply as storage units, holding substances within the cell until the contents can be used.
FIGURE 3-14 Vaults. Vaults are recently discovered organelles. They resemble tiny hollow pods and are thought to transport small structures to and from the nucleus. They can attach themselves to microtubules and rapidly zip from one part of the cell to another. The ends of each vault open to allow it to pick up or drop off loads.
NUCLEUS
The nucleus is the largest organelle in the cell and is considered the control center, the central processing unit, the CEO of operations, or the “brain” of the cell. It is a dominating, dark-staining, ovoid, spherical, flattened, or multisegmented body. The primary functions of the nucleus are to maintain the hereditary information of the species and to control cellular activities through protein synthesis. Thus the nucleus contains the hereditary information (DNA) that enables the cell to divide and produce an identical daughter cell and, on a larger scale, determines whether an animal will develop into a dog, cat, or horse. It also contains all of the instructions, blueprints, and information required to make over 2000 proteins that are needed for normal cell activity.
Although most cells have at least one nucleus, extremely large cells, such as muscle cells, may have many nuclei and are therefore called multinucleated. Mature mammalian red blood cells, on the other hand, have no nuclei, because the nuclei are removed from the cells during their development in the bone marrow. These cells are therefore called anucle- ate. Without a nucleus, they cannot divide, make protein or enzymes, or repair themselves as they start to age. For this reason, the supply of vital molecules in mammalian red blood cells allows them to survive in circulation for only 3 or 4 months. The red blood cells found in birds and reptiles, on the other hand, are nucleated and therefore are able to produce the proteins and other molecules needed by the cells to survive for longer periods.
The anatomy of the nucleus is divided into the following four parts:
• Nuclear envelope or membrane
• Nucleoplasm
• Chromatin
• Nucleoli
NUCLEAR ENVELOPE AND NUCLEOPLASM
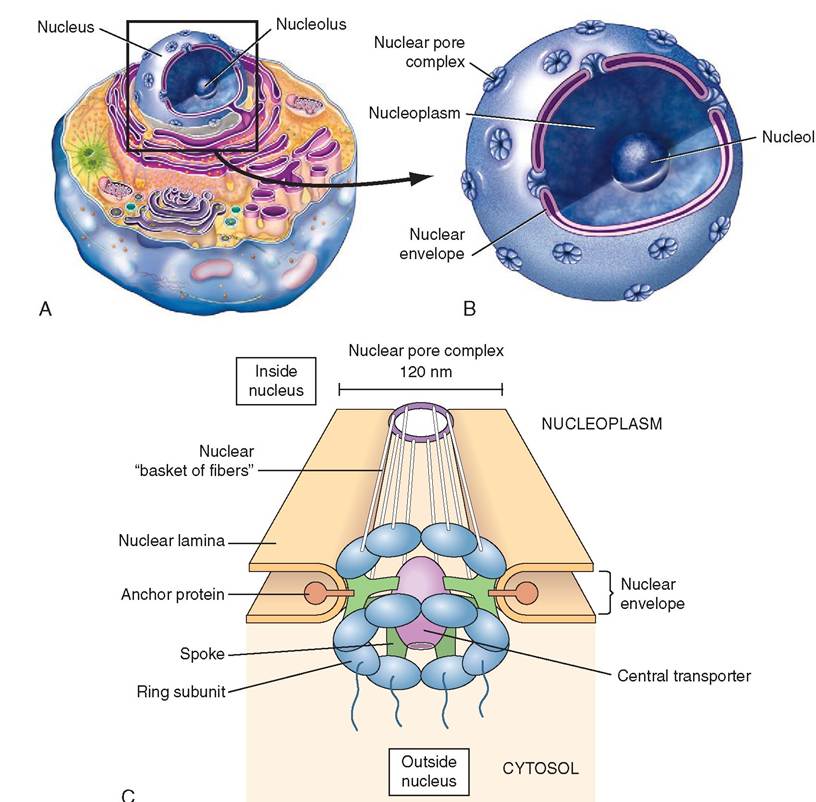
The nucleus is separated from the cytosol by a nuclear envelope composed of a lipid bilayer. The outer layer is continuous with the ER and is studded with ribosomes. Over 10% of the nuclear surface consists of nuclear pore complexes— places where the two layers of the nuclear envelope have fused to form a channel that spans its entire thickness (Figure 3-15). With improved microscopy techniques, the structure of nuclear pore complexes has become better understood, and they are relatively large (120 nanometers in diameter). Typically, protein molecules are moved into the nucleus from the cytoplasm, and mRNA and rRNA molecules are exported. The nuclear pore complexes represent the principal channels of communication between the cytoplasm and the nucleus. Between the bilayers of the nuclear envelope is a space called the perinuclear cisterna. The nucleus is filled with a gel-like substance called nucleoplasm that resembles cytosol and which contains chromatin.
DNA, RNA, AND CHROMATIN
The structure of genetic material, which was known to exist within the nucleus, was a mystery until the early 1950s, when research on heredity was to take a great leap forward. Rosalind Franklin (Box 3-2), a British physical chemist working at Kings College in London, used x-ray crystallography techniques to examine the structure of DNA. In 1953, she discovered that there are two forms of DNA, which she called A and B. It was her photograph 51 of the B form of DNA that showed the helical and double coaxial structure of the molecule, better known as the double helix. This critical discovery became the basis for the model that was subsequently developed by James Watson and Francis Crick (Figure 3-16) for which they won the Nobel Prize for Medicine and Physiology in 1962. Today, with the aid of advanced technology, cell biologists have an even better understanding of nucleic acids—DNA and RNA—and of the proteins that are central to the continuation of life.
DNA and RNA are made up of chains of nucleotides. Nucleotides are composed of three subunits: a nitrogenous base, a five-carbon sugar, and a phosphate group. In DNA the sugar is deoxyribose and in RNA the sugar is ribose. DNA and RNA nucleotides are linked in such a way as to form a “backbone” of alternating sugar and phosphate groups. The nitrogenous bases project out of this backbone and in DNA they are weakly bonded to nitrogenous bases on an opposing sntrand. I this way, DNA forms a double-stranded molecule, the basic structure of which is analogous to a twisted ladder in which the vertical poles are composed of alternating sugar and phosphate groups and the horizontal rungs are paired nitrogenous bases. DNA's molecular structure is therefore called the double helix (see Figure 3-16). RNA, however, is a single-stranded molecule that has no opposing strand. The single strand of RNA is similar in structure to each of the strands found in DNA.
Four kinds of nitrogen base are found in DNA and RNA nucleotides (Figure 3-17). The three that are found in both RNA and DNA are adenine (A), cytosine (C), and guanine (G). However, only DNA contains thymine (T), and only RNA contains uracil (U). In addition, the structure of each nitrogenous base permits the bonding of only certain pairs of nucleotides. For example, thymine can only bond to adenine, and cytosine can only bond to guanine. Uracil, the RNA base, can only bond to the DNA base adenine. These nitrogenous bases and their corresponding bonding parameters are the foundation for the storage of genetic information.
Chromatin appears as light or dark fibers in the nucleoplasm of the nucleus and is made up of DNA and globular proteins called histones. A single strand of DNA winds around eight histone molecules, forming a granule called a nucleosome. The nucleosomes are held together by short
us
FIGURE 3-15 Nucleus. A, The nucleus contains a nonmembrane bound region containing rRNA, called the nucleolus, and colloida nucleoplasm that contains chromatin. The entire nucleus is surrounded by a nuclear envelope that is perforated by tf^^L^s^^ls of nuclear pore complexes (NPC). The NPCs have a rosette appearance comprising eight 'petals' arranged eround a central transporter 'pore.' The NPC permits selective passage of substances in and out of the nucleus. B, A computer-enhanced transmission electron micrograph shows a nuclear pore and the details of the two lipid bilayer membranes that make up the nuclear envelope. Keep in mind that the outer lipid bilayer is continuous with the endoplasmic reticulum. Nucleoplasm (gray) indicle ⅛e nucleus and cytoplasm (yellow) outside the nucleus are also visible. C, A nud⅛ar pore complex is Comeosed of many parts. (A, Moefed fem Patton KT, Thibodeau GA: Anatomy & physiology, ed 8, S Louis, 201 3, Mosby. B, From Tliibodeau GA, Patton KT: Anatomy & physiology, ed 5, St Louis, 2003, Mosby.)
BOX 3-2 Rosalind Elsie Franklin
In the early 1950s, when research on heredity was moving forward rapidly, Dr. Rosalind Franklin, at King's College in Cambridge University, used crystal chromatography to study the structure of DNA. She made meticulous preparations and produced several excellent photographs. One in particular, photograph 51, showed the helical structure of the molecule. Without her permission, Maurice Wilkins, who worked with Franklin in the same laboratory, showed her data and photograph 51 to James Watson and Francis Crick. Immediately, Watson and Crick began work on a new 3-dimensional model that included two helical strands, discarding their previous 3-helix model. On March 17th, 1953, Franklin wrote a draft paper based on her own research describing the structure of DNA and detailed the placement of the sugars, phosphates and nitrogenous bases. However, before her paper was published, James Watson and Francis Crick's paper appeared on April 25, 1953 in Nature.
By the time Watson and Crick's paper was published, Franklin had left King's College for a new appointment at Birkbeck College, where she was given charge of a new research group working on the tobacco mosaic virus. At Birk- beck, Franklin found a collaborative working environment in which she became highly productive. She enthusiastically initiated work on the structure and assembly of the tobacco mosaic
BOX 3-2∣ Rosalind Elsie Franklin — confd

Rosalind Elsie Franklin. (From Piper A: Light on a dark lady. Trends Biochem Sci 23:151-154, 1998.)
virus and, during the four and a half years she spent at Birkbeck, published 17 papers. This was an impressive feat in the best of circumstances, but was particularly so for Franklin, because in 1956 she was diagnosed with ovarian cancer, which may have been caused by her lengthy exposures to x-ray radiation. Despite her illness, she maintained an arduous schedule and continued working in the laboratory until shortly before her death two years later. She died in London on April 16th, 1958 at the age of 37. After her death, her colleague Aaron Klug, who had been working with Franklin since 1954, took over the directorship of the laboratory at Birkbeck. He continued the research that Franklin had started on the tobacco mosaic virus and completed new research on the polio virus. Klug was subsequently awarded the Nobel Prize for this work.
In 1 962, four years after Rosalind Franklin's death, the Nobel Prize for Medicine or Physiology was awarded to Francis Crick, James Watson, and Maurice Wilkins for determining the structure of DNA. Regrettably, the Nobel Prize is only awarded to the loivmininga. tiNons cannot be made posthumously. The prema
ture death of Rosalind Franklin arguably robbed her of two cohances t win the Nobel Prize herself. Nevertheless, it is widely fheearltt t h contribution to the discovery of the structure of DNA iosmcparable to those who did receive the prize, and that she inmsdoienegd a the most brilliant scientists of our time.
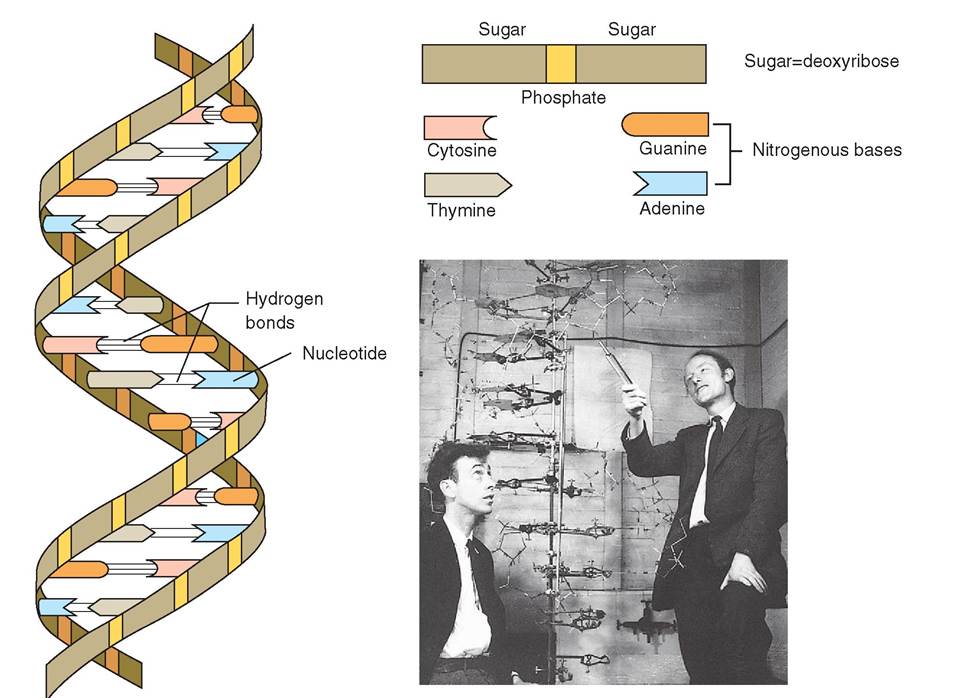
FIGURE 3-16 DNA. With the aid of data collected by Rosalind Franklin, of Britain, James Watson, of the United States, and Francis Crick, of Britain (photo, right), developed the double-helix model of deoxyribonucleic acid (DNA). Like a spiraling ladder, the vertical portion is composed of alternating molecules of sugar and phosphate, while the 'rungs' are paired nitrogenous bases. Watson, Crick, and Maurice Wilkins were awarded the Nobel Prize in 1962 for this famous work. (Photo from Cold Spring Harbor Laboratory.)
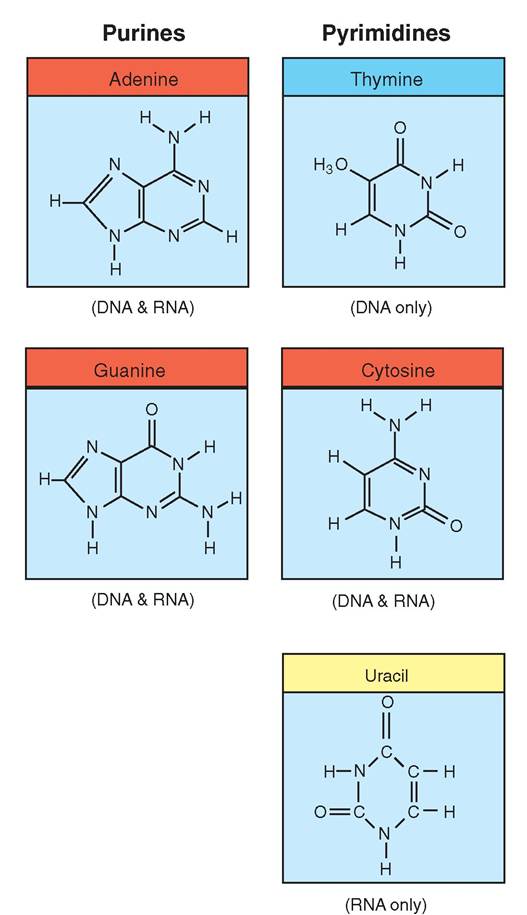
FIGURE 3-17 Nitrogenous bases. DNA and RNA are composed of two types of nitrogenous base: purines and pyrimidines. Purine molecules each have two rings and are found in both DNA and RNA. Pyrimidines, in contrast, are single-ringed molecules. Cytosine is the only pyrimidine that occurs in both RNA and DNA. Thymine occurs only in DNA, and uracil occurs only in RNA.
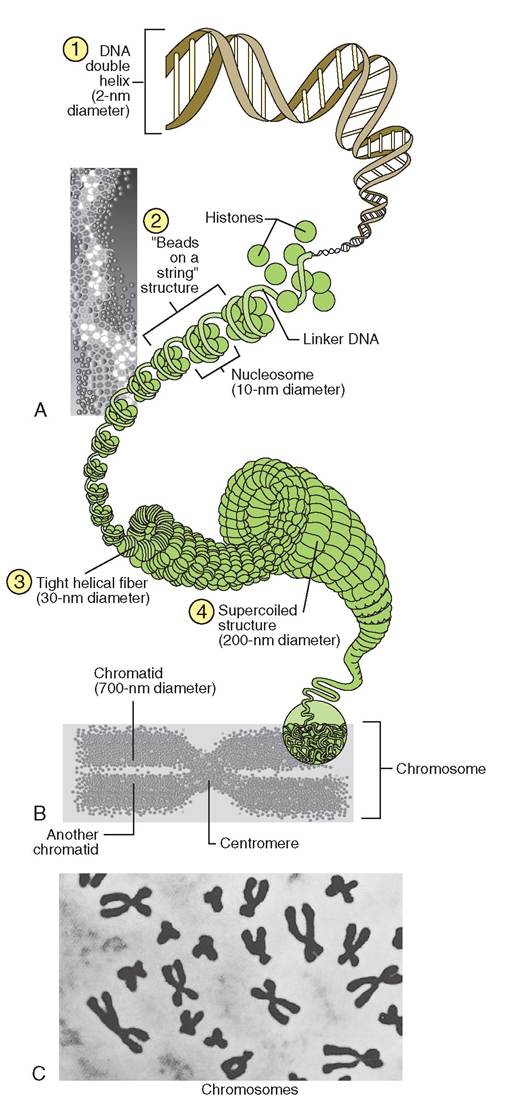
FIGURE 3-18 Chromatin (DNA and protein) comprises chromosomes. A, The double helix of DNA (1) wraps twice around eight histone proteins, forming a nucleosome. The nucleosomes are joined together by linker-DNA strands (2). Nucleosomes compress into a fiber, which coils tightly (3). The coiled fiber is then supercoiled and compressed into chromosomes (4). B, During cell division, the chromosome becomes shaped like an X, representing two chromatids held together at the centromere. C, Light micrograph of chromosomes in a cell preparing to divide. (C, From Thibodeau GA, Patton KT: Anatomy & physiology, ed 5, St Louis, 2003, Mosby.)
strands of DNA called linker DNA. Not only do the histone proteins help keep the DNA strand organized and untangled, they also expose small sections of the DNA to the outside nucleoplasm. These sections of DNA are called genes. By changing shape, the histones can expose different genes at different ⅛es. The exposed genes determine what proteins will be made by the cell. In this way, histones play an important role in the control of gene expression. We call this process gene regulation.
TAhe DN contains all of the important instructions required for the synthesis of thousands of different proteins, but not all of them are made: only a small percentage are
actually manufactured. The histones help to determine which segments of the DNA will be expressed and therefore which proteins will be made. When not being used to make protein, chromatin coils into tight helical fibers, which appear as dark strands in the nucleoplasm. This arrangement protects the delicate strands of DNA when they are not being used and saves space within the nucleoplasm. Strands of chromatin that are actively engaged in protein synthesis are uncoiled and called extended chromatin. The extended chromatin is lighter and is usually not visible under light microscopy. During cell division the chromatin condenses into supercoiled, X-shaped structures called chromosomes (Figure 3-18).
NUCLEOLI
Nuclei usually contain one or more small, dark-staining spherical patches known as nucleoli. The nucleoli are not membrane bound but are the places in the nucleus where ribosomal subunits are made. These subunits are exported separately from the nucleus and are assembled in the cytoplasm to form functional ribosomes. In addition, nucleoli contain the DNA that governs the synthesis of ribosomal RNA (rRNA).
TEST YOURSELF 3-4
1. Why do inclusions vary in appearance? What function do they perform?
2. What role does the centriole play in the formation of cilia and flagella?
3. How are centrioles structurally similar to cilia and flagella?
4. Why is the nucleus considered the "CEO of operations”?
5. Can a cell that does not contain a nucleus live as long as a cell that does contain one? Why or why not?
6. Describe the nuclear envelope. How is it different from the cell membrane?
7.How do histones play a role in gene regulation?
8. What is the significance of the nucleolus? What happens in that region of the nucleus?