BEHAVIORAL, PHYSIOLOGIC, AND ANATOMIC FEATURES
Behavioral Features
Adult male rabbits are called “bucks,” females “does,” and infants “kits.” Bucks must be kept separately, as they will fight to the death. Aggressive bucks may also traumatize does when breeding.
Does are induced ovulators, and do not have overt estrus cycles, although they have periodic intervals of receptivity. These periods can be detected by reddened vulvas, and conception rate is highest during this time. Following parturition (kindling), does leave their nest and return to nurse their altricial kits only once or twice daily. Prior to parturition, does create nests lined with fur plucked from their dewlaps, causing prepartum alopecia. Does can conceive almost immediately following kindling. Pseudopregnancy is common, and can be induced by a variety of stimuli, including mounting by other does, sterile matings by bucks, or bucks housed nearby. Female rabbits may be aggressive to unrelated litters of kits. Does, particularly nervous primiparous does, are prone to infant cannibalism. Rabbits engage in cecotrophy, or reingestion of soft mucus-coated “night feces.” Ceco- trophs are relatively high in protein and B complex vitamins. Rabbits are frequently nervous and easily frightened. When alarmed, rabbits may freeze (tonic immobility), run to shelter, or thump their hind feet as a group warning. Careful, firm handling, including support of the hind legs, is essential to avoid unrestrained kicking, which may result in accidental fracture of the vertebral column. Confined rabbits may exhibit various patterns of stereotypic behavior, barbering, obesity, and compulsive self-mutilation, particularly of digits. Rabbits mark their territory with urine, feces, scent glands, and by rubbing their chins (mental glands) on surfaces. The primary means of thermoregulation is through countercurrent blood exchange of ear vessels, panting, and salivation, all of which are inefficient. Thus, heat stroke is a significant problem for rabbits in hot climates. Wild rabbits effectively thermoregulate by living in burrows (warrens).Pathology of Laboratory Rodents and Rabbits, Fourth Edition. Stephen W. Barthold, Stephen M. Griffey, and Dean H. Percy. © 2016 John Wiley & Sons, Inc. Published 2016 by John Wiley & Sons, Inc.
Physiologic Features
Serum calcium levels up to 16 mg/dl are normal in rabbits. In contrast to most mammals in which intestinal calcium absorption is regulated according to metabolic needs, calcium is absorbed by rabbits in proportion to the amount in the diet, and intestinal uptake is not influenced by vitamin D. Serum calcium regulation is achieved through renal excretion. In rabbits, the renal fractional calcium excretion is 45%, compared to 2% in most mammals. Renal disease can, therefore, directly result in hypercalcemia, as well as through secondary hyperparathyroidism. Rabbits are very sensitive to hypercholesterolemia, since they are unable to increase sterol excretion in response to excess cholesterol in their diet. They exhibit hypercholesterolemia within days of being fed high-cholesterol diets, which has contributed to the use of rabbits for atherosclerosis research.
The rabbit gastrointestinal system is unique among herbivores. Their system is designed for maximal utilization of protein and vitamin-rich products of cecal fermentation while differentially eliminating indigestible fibrous waste. This is accomplished by a complex process that allows elimination of fibrous feces and reingestion of nutrient-rich cecotrophs. During the digestive process, fine digestible materials are retropulsed from the upper colon back into the cecum, while larger indigestible material is directed into the colon and passed as feces. This is facilitated by the spiral valve of the cecum, the taeniae of the haustrated upper colon, and the fusus coli, a ganglionic structure that separates the upper and lower colon. The fusus coli is known as the “pacemaker” of the rabbit intestine, and orchestrates the complex motility of the hind gut that is critical to cecotrophy.
This separation of digestible from nondigestible material facilitates the act of cecotrophy. Soft, mucous-covered cecotrophs (night feces) are selectively passed from the cecum to the anus during the act of coprophagy. Cecotrophs are major sources of B vitamins and protein, which are absorbed in the small intestine. Their mucous coating protects them as they pass through the acidic environment of the stomach.The major driving force of the rabbit digestive process is the presence of indigestible fiber, and lack thereof is a common cause of gastrointestinal disorders in rabbits. Intensive commercial production of rabbits has contributed to many of the gastrointestinal disorders in them. High-carbohydrate/low-fiber diets are deleterious to the digestive process. Pelleted diets should be supplemented with a source of fiber (hay). Perturbation of the microbiome by dietary and husbandry practices arises in a number of conditions, which precipitate severe and difficult to reverse dysbiosis. A stable enteric microbiome is essential to the health of the rabbit. Enteric microflora are minimal in the small intestine. The large bowel is populated by strict anaerobic bacteria, particularly Bacteroides spp. and facultative anaerobic bacteria such as Streptococcus spp. and Clostridium spp. are sometimes present in low numbers. Suckling and weanling rabbits are particularly prone to gastrointestinal disease. The cecal microbiota evolves from a simple and unstable microbiome after birth into a complex and stable community in sub-adult rabbits. The microbiome is a critical determinant of susceptibility to pathogens. For example, colonization of the ileum with segmented filamentous bacteria significantly inhibits colonization by enter- opathogenic Escherichia coli. An understanding of rabbit gastrointestinal disease is best approached with an understanding of rabbit gastrointestinal physiology. An excellent review of rabbit gastrointestinal physiology is available (see R.
& J.A.E Rees Davies).Anatomic Features External Features
Rabbits have 5 toes on their front feet and 4 toes on their rear feet, with dense fur on their rear foot pads. They may shed the fur 3 or 4 times a year, with hair growth cycles generally starting at the ventrum, growing dorsally and posteriorly, but regrowth may occur in irregular patchy configurations, which are obvious when the fur is clipped (Fig. 6.1). Rabbits develop folds of skin beneath their chins, known as dewlaps, which are especially prominent in mature does. The ear represents approximately 12% of the body surface area in domestic rabbits. Their size and accessible vessels make them ideal for venipuncture from the lateral veins, or collecting arterial blood from the central artery. Rabbits have inguinal and anal scent glands, as well as mental (chin) scent glands for territorial marking.
FIG. 6.1. Hair growth cycles generally start at the ventrum, progressing dorsally and posteriorly, but regrowth may often occur in irregular patchy configurations following clipping of the fur.
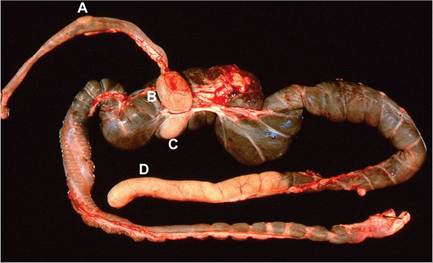
FIG. 6.2. Alimentary tract from a mature rabbit, illustrating the prominent gut-associated lymphoid tissue (GALT), including a Peyer's patch (A) in the wall of the ileum, the sacculus rotundus (B) at the terminal ileum, the cecal tonsil (C) in wall of the proximal cecum, and the cecal appendix (D) at the terminus of the cecum.
Central Nervous System
An incidental finding in normal rabbits is the presence of adipocytes in the interstitium of the choroid plexus.
HematologylClInIcal Chemistry
Erythrocytes are approximately 6.5-7.5 μm in diameter, and tend to be smaller in young rabbits. Polychromasia and anisocytosis are common, with 2-5% reticulocyto- sis. The mean life span for erythrocytes is relatively short, ranging from 57 to 67 days.
The counterpart of the neutrophil in rabbits is termed the heterophil. Heterophils are 9-15 μm in diameter and have distinct, acidophilic cytoplasmic granules. Experts have suggested that the neutrophil name should be retained in rabbits, since the characteristics of this cell resemble neutrophils more closely than the true heterophils that are seen in avian species. Eosinophils are 12-16 μm in diameter, with large cytoplasmic granules that stain a dull pink-orange with conventional hematology stains. Basophils may be relatively numerous and occasionally represent up to 30% of circulating leukocytes. Lymphocytes are usually the predominant leukocyte in the peripheral blood. Small lymphocytes are approximately 7-10 μm in diameter; large lymphocytes vary from 10 to 15 μm in diameter and may have a few azurophilic cytoplasmic granules. Normal hemogram and clinical chemistry ranges are published elsewhere. However, it is notable that rabbits tend not to develop leukocytosis, even with chronic bacterial infections.LymphoId System
Rabbits have a unique immunoglobulin repertoire. There is no evidence of IgD in rabbits. They possess a single IgG class that can activate complement as well as bind to Fcγ receptors, and the rabbit germline encodes 13 IgA isotypes, 2 of which are not expressed because of defective promoter regions. The thymus does not regress with age. Development of the primary antibody repertoire of rabbits occurs between 4 and 8 weeks of age, during which the VDJ genes of nearly all B cells undergo somatic diversification within the gut associated lymphoid tissue (GALT). Thus, rabbit GALT functions in a manner similar to the Bursa of Fabricius in birds. In addition, this process is dependent upon select species of normal intestinal microflora, which drive gene diversification in a nonantigen-dependent manner.
The GALT represents over 50% of the total mass of lymphoid tissue in the body, which accounts for the relatively small spleen in this species.
The GALT includes Peyer's patches of the small intestine, as well as structures that are unique to the rabbit. The sacculus rotundus is a spherical, thick-walled enlargement of the terminal ileum at the ileocecal junction (Fig. 6.2). The adjacent cecum has a round patch of lymphoid tissue called the cecal tonsil. The tip of the cecum contains the prominent thick-walled cecal appendix. These lymphoid structures contain aggregates of organized lymphoid tissue in the lamina propria and submucosa. A common incidental finding in rabbits is the presence of large histiocytes filled with particulate debris in the follicular centers of the GALT (Fig. 6.3).Transfer of maternal immunoglobulins from dam to kit is mediated predominantly in utero through yolk sac receptors, although postnatal transfer also occurs from birth to 12 days of age. Passively transferred immunoglobulin begins to wane around 3 weeks of age.
CardIovascular System
The right chambers of the heart are relatively thinwalled, and a frequent postmortem finding is a quantity of clotted blood in the right ventricle, with no evidence of postmortem contraction. The right atrioventricular valve, which is tricuspid in most mammals, is bicuspid in rabbits.
Respiratory System
Rabbits are obligate nasal breathers, and exhibit marked dyspnea when nasal passages are obstructed. The rabbit lung does not contain respiratory bronchioles. Airways
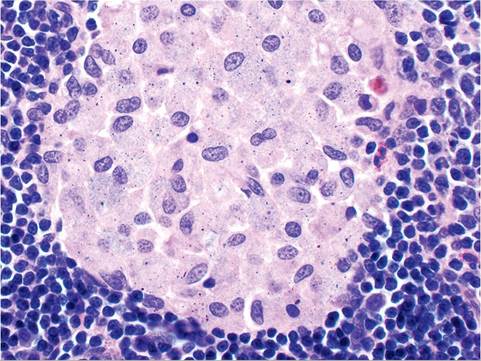
FIG. 6.3. GALT from a normal mature rabbit, depicting the common presence Ofhistiocytes filled with refractile particulate debris within the follicular center.
terminate in vestibules that contain alveoli. Pulmonary arteries are enveloped in a prominent smooth muscle layer, which can be misinterpreted as hypertrophy (Fig. 6.4).
Gastrointestinal System
Rabbits have a dental formula of I2/1, C0/0, P3/2, M3/3, with continuously growing (elodont) incisors as well as cheek teeth. Lagomorphs have 4 upper incisors, including peg teeth aligned directly behind the upper front incisors. Rarely, rabbits may congenitally lack peg teeth, but can function well as long as there is not malocclusion. Although not generally noticed, rabbits have diphyodont dentition (deciduous and permanent teeth). Deciduous teeth are shed around birth, including a third pair of incisors, lateral to the upper incisors. As
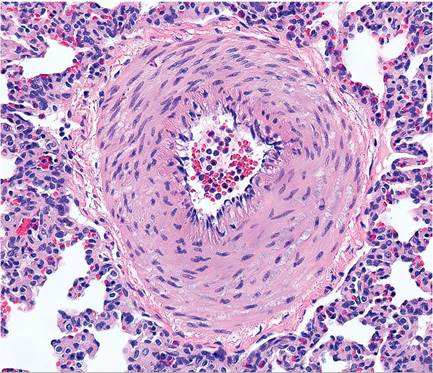
FIG. 6.4. Pulmonary artery of a normal rabbit. Pulmonary arteries are highly muscular, which can be misinterpreted as muscular hypertrophy.
with other herbivores, rabbits have a large and relatively complex digestive system, which is a challenge to dissect at necropsy. The stomach is thin-walled and typically contains approximately 15% of the ingesta present in the alimentary tract. The small intestine is short compared to that in many species, representing roughly 12% of the total volume of the gastrointestinal tract. Brunner's glands are distributed throughout the length of the duodenum. The bile and pancreatic ducts have separate openings into the duodenum. The cecum typically holds around 40% of the ingesta present in the digestive system. The colon consists of 2 major segments, the craniad portion being haustrated with prominent muscular taeniae. The mucosal surface of the proximal upper colon has multiple wart-like projections (warzen) that increase surface area. The lower colon lacks haustra and taeniae. These regions of the colon are divided by a region known as the fusus coli, which is 4-5 times thicker than the lower colonic wall, and contains aggregates of ganglia, which are involved in the highly coordinated digestive process.
The intestinal mucosa is endowed with copious numbers of goblet cells, which occupy the depths of crypts to the tips of villi. This is often misdiagnosed as goblet cell hyperplasia. In the liver, hepatocytic vacuolation associated with glycogen accumulation is a variable finding in rabbits fed with commercial diets. As in rodents, aged rabbit livers may feature polykarya, anisokarya, and intranuclear invagination of cytoplasm. The rabbit liver secretes biliverdin, rather than papilla, and rabbits produce copious amounts of bile compared to other species. The pancreas is a poorly defined collection of lobules adjacent to the duodenum.
Urogenital System
Once rabbits begin eating solid rations, the alkaline urine normally contains large quantities of dull yellow to brown ammonium magnesium phosphate and calcium carbonate monohydrate crystals. Occasionally, normal rabbit urine may be dark red to orange, which is due to porphyrins. Hyperpigmented urine has also been associated with elevated levels of urobilin, the oxidative product of urobilinogen. Pigmented urine must be differentiated from hematuria, which may arise due to uterine adenocarcinoma, uterine polyps, endometrial venous aneurysms, cystitis, urinary bladder polyps or tumors, pyelonephritis, and renal infarction with hemorrhage. Rabbit kidneys have a single papilla, and ectopic glomeruli can be frequently found in the medulla and peripelvic regions.
The rabbit has a bicornuate uterus comprised of 2 separate uterine horns and 2 cervices (Fig. 6.5). Placentation is hemochorial and bidiscoidal. Neonatal rabbits receive most of their maternal antibody via transplacental transfer during pregnancy. Both anterior and breach deliveries are normal. The testes of bucks descend at around 12 weeks of age, but inguinal canals remain
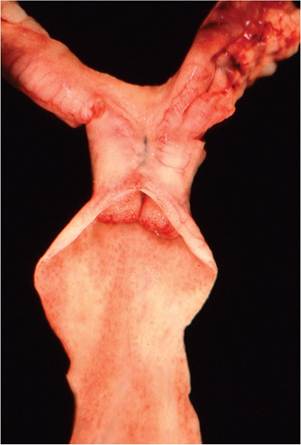
FIG. 6.5. Female reproductive tract of a normal doe, depicting the 2 separate cervices, which is typical for this species. The uterus is bicornuate, with 2 separate horns.
open and testes can be retracted abdominally. Scrotums are hairless. Testes may undergo seasonal involution, which may involve abdominal retraction, with testicular size reduced up to one-half their normal size. During this phase, spermatid giant cells and atrophic changes may be apparent microscopically. Seasonal infertility may occur in bucks in hot climates.
Musculoskeletal System
In the domestic rabbit, bones are relatively fragile. The skeleton of New Zealand White rabbits represents approximately 6-7% of the total body weight. On the other hand, skeletal muscle constitutes over 50% of the body weight in this species. Fractures, particularly those of the vertebral column, readily occur, particularly when the hind legs are not restrained properly during handling.
BIBLIOGRAPHY FOR BEHAVIORAL, PHYSIOLOGIC, AND ANATOMIC FEATURES
Carneiro, M., Alfonso, S., Geraldes, A., Garreau, H., Bolet, G., Boucher, S., Tircazes, A., Queney, G., Nachman, M., & Ferrand, N. (2011) The genetic structure of domestic rabbits. Molecular Biology and Evolution 28:1801-1816.
Christensen, N.D. & Peng, X. (2012) Rabbit genetics and transgenic models. In: The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents (eds. M.A. Suckow, K.A. Stevens, & R.P. Wilson), pp. 165-193. Academic Press, London.
Clark, M.R. (1997) IgG effector mechanisms. Chemical Immunology 65:88-110.
Clauss, M., Burger, B., Liesegang, A., Del Chicca, F., Kaufmann-Bart, M., Riond, B., Hassig, M., & Hatt, J.-M. (2011) Influence of diet on calcium metabolism, tissue calcification and urinary sludge in rabbits (Oryctolagus cuniculus). Journal of Animal Physiology and Animal Nutrition 96:798-807.
Combes, S., Michelland, R.J., Monteils, V., Cauquil, L., Soulie, V., Tran, N.U., Gidenne, T., & Fortun-Lamothe, L. (2011) Postnatal development of the rabbit caecal microbiota composition and activity. FEMS Microbiology and Ecology 77:680-689.
Crossley, D.A. (1995) Clinical aspects of lagomorph dental anatomy: The rabbit (Oryctolagus cuniculus). Journal of Veterinary Dentistry 12:137-140.
Heczko, U., Abe, A., & Finlay, B.B. (2000) Segmented filamentous bacteria prevent colonization of enteropathogenic Escherichia coli 0103 in rabbits. Journal of Infectious Diseases 181:1027-1033.
Mage, R. G., Lanning, D., & Knight, K.L. (2006) B cell and antibody repertoire development in rabbits: the requirement of gut-associated lymphoid tissues. Developmental and Comparative Immunology 30:137-153. MediRabbit.com
Naff, K.A. & Craig, S. (2012) The domestic rabbit, Oryctolagus cuniculus: origins and history. In: The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents (eds. M.A. Suckow, K.A. Stevens, & R.P. Wilson), pp. 157-163. Academic Press, London.
Peri, B.A. & Rothberg, R.M. (1996) Transmission of maternal antibody prenatally and from milk into serum of neonatal rabbits. Immunology 57:49-53.
Rees Davies, R. & Rees Davies, J.A.E. (2003) Rabbit gastrointestinal physiology. Veterinary Clinics of North America Exotic Animal Practice 6:139-153.
Sohn, J. & Couto, M.A. (2012) Anatomy, physiology, and behavior. In: The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents (eds. M.A. Suckow, K.A. Stevens, & R.P. Wilson), pp. 195-215. Academic Press, New York.
Suckow, M.A., Brammer, D.W., Rush, H.G., & Chrisp, C. (2002) Biology and diseases of rabbits. In: Laboratory Animal Medicine, 2nd edn (eds. J.G. Fox, L.C. Anderson, F.M. Loew, & F.W. Quimby), pp. 329-364. Academic Press, New York.
Suckow, M.A. & Schroeder, V. (2010) The Laboratory Rabbit, Laboratory Animal Pocket References. CRC Press, Boca Raton, FL.
Tsunenari, I. & Kast, A. (1992) Developmental and regressive changes in the testes of the Himalayan rabbit. Laboratory Animals 26:167-179.
Washington, I.M. & Van Hoosier, G. (2012) Clinical biochemistry and hematology. In: The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents (eds. M.A. Suckow, K.A. Stevens, & R.P. Wilson), pp. 57-116. Academic Press, London.
Wells, M.Y., Weisbrode, S.E., Maurer, J.K., Capen, C.C., & Bruce, R.D. (1988) Variable hepatocellular vacuolization associated with glycogen in rabbits. Toxicologic Pathology 16:360-365.